Возбудитель болезни, Фитопатоген

Возбудитель болезни растений (фитопатоген) – патогенный организм, вызывающий болезнь растения [2] .
Ранее понятие «возбудитель болезни растений», кроме патогенных организмов включало прочие экологические факторы, обуславливающие возникновение болезни растений, в том числе: насекомые, метеорологические факторы, токсические вещества, недостаток или избыток питательных веществ [5] .
Содержание:
Найти в природе растение без посторонней микробиоты невозможно. Многие здоровые растения содержат возбудителей болезни в латентной форме. Несмотря на этот неопровержимый факт, подавляющее большинство растений здоровы, то есть все основные биохимические и биофизические процессы проходят в определенных, среднестатистических рамках. Под влиянием возбудителя заболевания или неблагоприятных абиотических факторов окружающей среды наблюдается нарушения нормального развития – болезнь растения [1] .

Корневая гниль сеянцев огурца

Возбудители болезни – Pythium debarianum, Pythium spinosum, Pythium spinosum.
1 – Сеянцы огурца, пораженные ризоктониозной корневой гнилью (возбудитель Rhizoctonia solani) [1] ; 2 – Гифы мицелия грибов рода Pythium [1] ; 3 – Зооспорангии Pythium debarianum [1] ; 4 – Ооспоры Pythium spinosum [1] .
Систематические категории возбудителей болезни растений
Возбудителями болезни растений являются различные микроорганизмы:
Вирусы – неклеточные субмикроскопические организмы. Состоят из нуклеиновой кислоты, покрытой белковой оболочкой. Ведут паразитический образ жизни внутри живых клеток [1] .
Вироиды – вирусоподобные фитопатогены, состоящие из одной циклической молекулы РНК, способной реплицироваться в растении [1] .
Бактерии – одноклеточные безъядерные организмы (прокариоты). Нуклеотид бактериальной клетки состоит из кольцевой ДНК. Нуклеотид не отделен от цитоплазмы мембраной и имеет вид небольших зерен [1] .
Фитоплазмы – специфическая группа организмов, занимает промежуточное положение между бактериями и вирусами [1] .
Грибы и грибоподобные организмы – отдельная группа организмов, отличающаяся от растений и животных типом питания. В отличие от растений, они не являются автотрофами и утилизируют только запасенную в органических соединениях энергию. В отличие от животных грибы – осмотрофы. Питаются всасыванием вещества из окружающей среды [6] .

Представители систематических категории фитопатогенов

1 – Тела включения вируса мозаики томатов в ткани растения-хозяина садовый помидор [12] ; 2 – Культура бактерии Xanthomonas campestris, возбудителя сосудистого бактериоза крестоцветных культур [8] ; 3 – Гифы гриба Verticillium dahlia – возбудитель вертициллеза огурца [13] ; 4 – Грибоподобный огранизм, миксомицет, возбудитель порошистой парши картофеля – Spongospora subterranea [11] .
Свойства фитопатогена
Патогенность
Патогенность – специфическая способность микроорганизма вызывать заболевание [1] .
Агрессивность
Агрессивность – количественный признак патогенности, отражающий способность патогена к размножению в тканях растений, на которых он паразитирует. Агрессивность изменяется в широком диапазоне в зависимости от условий окружающей среды. Агрессивность оценивается в зависимости:
- от длины инкубационного периода – быстроты появления симптомов;
- скорости распространения патологического процесса по тканям растений;
- числа инфекционных единиц, способных вызывать заражение;
- от интенсивности спороношения (у грибов) [1] .
Вирулентность
Вирулентность – качественный признак патогенности, ее можно определить, как способность фитопатогена вызывать заболевания у определенных видов и сортов растения-хозяина [1] .
Установлено существование специализированных рас патогенов, которые вирулентны для одних сортов и невирулентны (авирулентны) для других [1] .
Специализированные формы распадаются на физиологические расы, а последние в свою очередь – на биотипы, отличающиеся по патогенности от основной расы [7] .
Физиологические расы – группы патогенов внутри вида, способные паразитировать только на определенных сортах растений-хозяев. Физиологические расы отличаются генотипами. Название «физиологические» подчеркивает, что различие между расами основано на различии в физиологии заражения. Они определяются генетически. Способность или неспособность расы вызывать заражение того или иного сорта связано с различными способами воздействия этих рас на растение, с различными физиологическими реакциями [7] .
На одном и том же сорте различные расы ведут себя по-разному: одна устанавливает взаимоотношения с клетками растения, получает от него питательные вещества, другая вызывает в клетках защитную реакцию сверхчувствительности, в результате которой наблюдается гибель и клетки, и патогена [7] .
Состав рас патогена отличается в различных регионах и определяется культивируемыми в данной местности сортами растения-хозяина [7] .
Например, возбудитель стеблевой (линейной) ржавчины зерновых, базидиальный гриб из Порядка Ржавчинные – Puccinia graminis, имеет более 300 рас. Внутри расы могут подразделяться на биотипы, отличающиеся по патогенности от основной расы. В США у расы 15 Puccinia graminis были выявлен биотип 15А, образующий более мелкие пустулы и 15В с более высокой вирулентностью на дополнительном сорте-дифференциаторе [7] .
Физиологические расы обозначаются цифрами, биотипы – буквой, стоящей после цифрового обозначения расы [7] .
Формировать физиологические расы и биотипы способны патогены, относящиеся к различным систематическим категориям фитопатогенов. Физиологические расы и биотипы установлены у слизевиков, грибов, цветковых паразитов. Вирусы и бактерии формируют аналогичные расам группы – штаммы [7] .
Специализация фитопатогенов
Специализация фитопатогенов – приуроченность к определенному питающему субстрату, способность паразитировать на одном или нескольких растениях-хозяевах [7] .
Различают несколько видов специализации:
- филогенетическая – проявляется в приспособленности паразитировать на близкородственной группе растений-хозяев;
- онтогенетическая (возрастная) – связана с приуроченностью патогенов к определенному этапу развития растения или отдельных органов;
- гистотропная – способность поражать определенные ткани растений;
- органотропная – способность поражать определенные органы растений.
Фитопатоген может обладать всеми типами специализации либо одним или несколькими. Наиболее важны филогенетическая и онтогенетическая специализации [7] .
Филогенетическая специализация
Филогенетическая специализация – способность патогенного вида к поражению одного или многих видов растений-хозяев. По данной характеристике патогены делят на полифагов и монофагов:
Инфекционные и вирусные болезни растений: особенности патогенеза

Вирусные и инфекционные заболевания сельскохозяйственных растений – постоянная «головная боль» тех, кто ухаживает за полезными культурами.
Вирусные и инфекционные заболевания сельскохозяйственных растений – постоянная «головная боль» тех, кто ухаживает за полезными культурами. Нисколько не меньше хлопот доставляют они и дачникам, ведь вирусы и прочие патогены обладают уникальной способностью в кратчайшие сроки приспосабливаться к изменяющимся внешним факторам и очень быстро адаптируются даже к самым новым препаратам.
Инфекционные, или паразитарные, болезни растений вызывают патогенные микроорганизмы (фитопатогены). Основной признак инфекций болезней — заразность, т. е. способность передаваться от растения к растению. Начинаются они, как правило, с единичных растений в агроценозе, а впоследствии могут приобретать массовый характер распространения. Развитие инфекции растений и ее интенсивность не являются постоянными для одной и той же местности, а колеблются от малозаметных случаев до массового поражения растений — эпифитотии. Для возникновения эпифитотий имеют большое значение способности патогена к накоплению инфекции, к образованию новых рас, скорость распространения инфекции, длительность сохранения жизнеспособности патогенного организма, а также устойчивость растений и условия окружающей среды.
Вирусы растений — это группа облигатных внутриклеточных паразитов, характеризующихся ультрамикроскопическими размерами, отсутствием клеточного строения и размножением только в клетках живых организмов. Первоначально открыты как возбудители болезней растений (мозаичная болезнь табака) в 1892 году русским ученым Д. И. Ивановским. Свое название эти патогены получили в 1898 году от голландского микробиолога М. Бейеринка, продолжившего исследования мозаики табака. Вирусы способны вызывать серьезные заболевания растений, приводящие к потере урожая и к ухудшению его качества; теряется сорточистота, холодостойкость, зимостойкость растений. Поражение вирусами, вызывающими заболевания растений, отрицательно влияет на пищевую и кормовую ценность продукции, пригодность ее к промышленной переработке. Особенно вредоносны они при выращивании семенного и посадочного материала. Вызываемое вирусами у растений явление стерильности и несовместимости отрицательно отражается на результатах работы селекционеров.
Патогенез инфекционных заболеваний
Различают местные эпифитотии, развивающиеся из года в год на определенной ограниченной территории (поле, сад, теплица). Возбудители их медленно накапливаются в природе (фузариозное и вертициллезное увядание многих культур). Прогрессирующие эпифитотии возникают на обширных территориях, но в сильной мере зависят от условий погоды. Часто отмечают прогрессирующие эпифитотии бурой, желтой и стеблевой ржавчины пшеницы. Для вирусных и фитоплазменных болезней определяющим фактором является массовое отрождение или миграция насекомых-переносчиков. Повсеместные эпифитотии (панфитотии) могут охватывать целые страны и континенты, но случается редко. К широко известным примерам повсеместных эпифитотий можно отнести вспышки фитофтороза картофеля в Европе в 40-х годах XIX века, ржавчины на зерновых на юге России в начале XX века.
Любой фитопатоген воздействует на клетки растения при помощи продуктов своего обмена веществ (токсинов, ферментов, физиологически активных веществ), забирает из них органические вещества и может распространяться по растению, нарушая нормальный процесс жизнедеятельности, т. е. вызывая патологический процесс. Этот процесс может возникнуть и развиваться только при наличии восприимчивого к определенному патогену растения-хозяина, достаточного количества инфекционного материала, контакта паразита и растения-хозяина в определенных условиях окружающей среды.
Все возбудители болезней растений обладают патогенностью, т. е. способностью вызывать заболевания растений, приводящие к снижению их продуктивности или гибели; вирулентностью — качественным признаком, который обозначает патогенность по отношению к отдельным культурам и сортам; агрессивностью — количественным признаком патогенности, отражающим интенсивность развития болезни (симптомов) на растении (сорте), варьирующимся в зависимости от условий окружающей среды.
Растение как среда обитания также оказывает определенное воздействие на возбудителя болезни. Восприимчивые растения не способны противостоять заражению и дальнейшему развитию патогена. Толерантные (выносливые) растения поражаются болезнью, возникает патологический процесс, но при этом продуктивность растений практически не снижается. Устойчивые сорта или виды растений поражаются болезнью в очень слабой степени и фактически не снижают урожайность. Иммунные к определенным болезням сорта вообще ими не поражаются.
Большую роль в возникновении и развитии болезни играют погодно-климатические условия, которые влияют как на самого возбудителя болезни, так и на растение-хозяина, ослабляя его, или, наоборот, повышая его комплексную устойчивость. Причем оказывают влияние не только колебания погоды в течение вегетационного сезона на обширной территории, но и изменения микроклимата на ограниченных площадях из-за различий рельефа, качества обработки почвы.
Выделяют три основных этапа патогенеза инфекционных заболеваний сельскохозяйственных растений:
- Заражение, или проникновение возбудителя в ткани растения. Большая часть патогенов проникает через естественные отверстия: устьица, чечевички, гидатоды, рыльце пестика (возбудители ложных мучнистых рос, многие бактерии). Другие патогены проникают через различные макро- и микротравмы тканей растений (возбудители монилиозной плодовой гнили, цитоспороза, черного рака плодовых, ряд вирусов и бактерий). Небольшая группа грибов может внедряться через неповрежденные покровные ткани растения (возбудители настоящих мучнистых рос). Большое значение для успешного проникновения патогена имеют выделения растений, их количественный и качественный составы, которые зависят от вида, сорта растения, его возраста, физиологического состояния.
- Развитие, или накопление биомассы. В этот период, попав на благоприятный субстрат, патогены начинают питаться и размножаться способом, зависящим от их биологических особенностей (делением, бесполым и половым размножением, саморепликацией). Заселение патогенов может быть локальным (пузырчатая головня кукурузы), а при системной реакции они распространяются по всему растению (пыльная и твердая головня пшеницы). В процессе взаимодействия растения и возбудителя болезни возникают общие физиологические и биохимические изменения, а вслед за ними — и анатомо-морфологические изменения.
- Появление симптомов, или визуализация болезни. На этом этапе в результате взаимодействия фитопатогена, растения-хозяина и факторов внешней среды происходят патологические изменения, характерные для данной болезни. Тип симптомов зависит в первую очередь от вида возбудителя. Для большинства грибных болезней окончательным симптомом является появление спороношения на поверхности растения.
В патологическом процессе период от проникновения патогена в растение до появления массовых симптомов заболевания растений называется инкубационным периодом. Его продолжительность зависит от биологических особенностей возбудителя (скорости его развития и распространения в тканях растения), степени восприимчивости растения и условий окружающей среды, особенно температуры. Например, у головневых грибов инкубационный период продолжается несколько месяцев, а у мучнисторосяных грибов — несколько дней. Инкубационный период возбудителя фитофтороза картофеля (Phytophthora infestans) при среднесуточной температуре 18-20 °С равен 4 дням, а при температуре 13-15 °С увеличивается до 10 дней. Влажность окружающей среды также может оказывать влияние на длину инкубационного периода, особенно при грибковых заболеваниях.


Для проведения краткосрочных прогнозов развития болезни важно знать продолжительность инкубационного периода и уметь правильно его рассчитать, это позволит своевременно провести, например, опрыскивание растений фунгицидами и предотвратить распространение болезни на другие участки.
Возбудители инфекций растений
Возбудители болезней растений (грибы и псевдогрибы, бактерии, фитоплазмы, вирусы, вироиды, цветковые растения-паразиты) весьма многочисленны, их описано свыше 40 000 видов. Они различаются по уровню организации, систематическому положению, типу паразитизма, специализации и другим признакам. Фитопатогенные организмы широко представлены во всех биоценозах благодаря таким биологическим особенностям, как скорость размножения, высокая биохимическая активность, генетическая изменчивость, адаптивность к неблагоприятным факторам среды, возможность длительно пребывать в состоянии анабиоза и переходить к активной жизнедеятельности при благоприятных условиях.
По степени паразитизма можно выделить три типа фитопатогенных организмов. Факультативные паразиты основную часть жизненного цикла питаются органическим веществом мертвых клеток растений — сапротрофно. Они первоначально поражают ослабленные, имеющие повреждения растения, заселяя вначале участки отмерших тканей. Это самая многочисленная группа возбудителей болезней, включающая все фитопатогенные бактерии и большинство грибов. Один из эффективных приемов защиты от этой группы патогенов — создание наиболее благоприятных условий для роста и развития растений, в результате чего они становятся менее подверженными заболеваниям.
Облигатные паразиты питаются органическим веществом живых клеток, развиваются только в живых тканях растений. К ним относятся все фитопатогенные вирусы и вироиды, грибы — возбудители ржавчины, ложных и настоящих мучнистых рос. Изменение условий внешней среды, как правило, влияет только на сроки появления и интенсивность заболевания. Использование устойчивых сортов и гибридов является радикальным способом борьбы с облигатными паразитами. Однако для эффективной защиты следует сочетать использование устойчивых сортов, средств защиты растений, подавляющих процессы накопления и распространения возбудителей в период вегетации, и агротехнических приемов, направленных на сокращение количества инфекционного начала.


Факультативные сапротрофы являются небольшой промежуточной группой, которая чаще питается органическим веществом живых клеток, быстро приводя их к гибели. К ним относятся Phytophthora infestans — возбудитель фитофтороза пасленовых, Venturia inaequalis — возбудитель парши яблони, Ascochyta pisi — возбудитель аскохитоза гороха.
Принято выделять несколько типов специализации возбудителей болезней. Филогенетическая специализация проявляется в приспособленности патогенов к питанию на растениях определенного семейства, рода, вида и даже сорта. Широко специализированные патогены, или полифаги, паразитируют на растениях разных семейств или внутри одного семейства на разных родах.


Так, возбудитель серой гнили Botrytis cinerea поражает землянику, огурец, капусту, морковь, фасоль и многие другие культуры. К узкоспециализированным возбудителям болезней, или монофагам, поражающим растения одного рода или вида, относится, например, гриб — возбудитель рака картофеля Synchytrium endobioticum, поражающий картофель. Патогены одного вида подразделяются на еще более специализированные формы — физиологические расы, различающиеся только по способности паразитировать на определенных сортах растения-хозяина. Онтогенетическая, или возрастно-физиологическая, специализация определяет способность патогена поражать растения, находящиеся в определенной фазе своего развития. Такая приуроченность связана с различным состоянием продуктов метаболизма растения и питательными свойствами его тканей в различные периоды роста и развития. Органотропная, или тканевая, специализация характеризуется приуроченностью патогенов к определенным органам и тканям растения. Для иллюстрации тканевой специализации можно привести
возбудителей настоящих мучнистых рос, питающихся в основном в тканях эпидермиса.
Для организации эффективной защиты от болезней необходимо знать, каким образом и где может сохраняться возбудитель конкретной болезни, а также — как он распространяется в период вегетации. В связи с этим выделяют понятия источников первичной и вторичной инфекции.
Источник первичной инфекции, или первичная инфекция, — это болезнетворное начало, которое впервые в данный вегетационный период после сохранения в неблагоприятных условиях способно вызвать заражение растений. У грибов она может быть представлена различными формами: склероция-ми, цистами, ооспорами, клейстотециями, телиоспорами и др. На практике под источником первичной инфекции понимают место (субстрат-носитель) сохранения патогена в неблагоприятный для него период. Это может быть почва, растительные остатки, семена и т. п. Хотя первичное заражение происходит обычно за счет перезимовавшей инфекции, она может появиться и с приобретенным посадочным материалом, семенами или извне, перенесясь в виде спор грибов с далекого расстояния. Зимующие стадии возбудителей болезней иногда бывают очень стойкими и могут сохраняться (например в почве) в течение нескольких лет (покоящиеся споры возбудителя килы капусты — Plasmodiophora brassicae). Длительность сохранения первичной инфекции следует учитывать при составлении севооборотов.
Источником вторичной инфекции называют болезнетворное начало, обеспечивающее перезаражение растений в течение вегетационного периода и основной способ его распространения. Вторичная инфекция у грибных патогенов может быть представлена различными формами: зооспорами, спорангиоспорами, конидиями, урединиоспорами, обрывками мицелия. Заражение растений некоторыми болезнями происходит только один раз за вегетационный период. Такие болезни называютмоноциклическими, вторичная инфекция у них отсутствует. К ним относятся, например, головневые болезни, курчавость листьев персика, красная пятнистость листьев сливы. При большинстве других заболеваний их называют полициклическими, после завершения инкубационного периода формируется инфекция (генерация), способная вызвать перезаражение других растений в этот же вегетационный период, причем происходит это неоднократно. Примеры полициклических болезней — американская мучнистая роса смородины и крыжовника, парша яблони (за вегетационный период образуют более 10 генераций конидий), корончатая ржавчина овса (образуется 2-3 генерации урединиоспор).
Способы сохранения патогенов. Возбудители болезней растений, в основном грибы и бактерии, могут сохраняться в растительных остатках (стерне, листьях, плодах, корнях и т. п.), находящихся на поверхности почвы или в неглубоком почвенном слое. В послеуборочных остатках обычно сохраняются бактерии; в форме ооспор в них зимуют многие возбудители ложных мучнистых рос. Почва также может служить резерватором и накопителем инфекции. Некоторые почвообитающие фитопатогенные грибы из родов Fusarium, Rhizoctonia, бактерии рода Agrobacterium могут жить в почве и вне пораженных растений, при благоприятных условиях переходя к паразитическому образу жизни. Однако большая часть патогенов не способна к длительному существованию в почве и лишь сохраняется в ней в форме покоящихся структур, таких, как склероции, хламидоспоры, цисты, ооспоры.


Почва является основным источником первичной инфекции для возбудителей склеротиниоза озимых зерновых, ложной мучнистой росы подсолнечника, корневых гнилей зерновых, рака картофеля. В почве сохраняются семена цветковых растений-паразитов — повилики, заразихи. Представители всех фитопатогенных микроорганизмов могут сохраняться в семенном и посадочном материале. Они могут быть в виде примесей (склероции грибов), на поверхности, например, семян (телиоспоры твердой головни пшеницы), внутри семян (мицелий пыльной головни пшеницы) и посадочного материала (вирусы и бактерии в клубнях картофеля). Пораженные многолетние растения (в т. ч. сорняки, озимые, тепличные культуры) при возобновлении болезни на следующий год становятся первичными источниками инфекции для здоровых растений. Таким способом сохраняются многие вирусы, фитоплазмы, бактерии, некоторые грибы. Дополнительными источниками сохранения инфекции могут быть насекомые и клещи-переносчики, а также другие организмы, в теле которых могут зимовать многие фитоплазмы и вирусы. Например, вирус полосатой мозаики пшеницы сохраняется в теле галообразующих клещей рода Eriophyes.
Пути распространения патогенов. Разнообразие путей распространения возбудителей болезней растений следует учитывать при прогнозе болезней, мерах профилактики и защиты, сдерживающих появление их в вегетационном сезоне. Наиболее часто встречается распространение воздушным путем (анемохория), с водой (гидрохория), с помощью различных организмов-переносчиков (зоохория) и человека (антропохория). Распространение ряда патогенов происходит при вегетативном размножении растений. Многие грибы, например возбудители ложных мучнистых рос, распространяются воздушно-капельным путем, т. е. одновременно с помощью воды и воздуха, только в таких условиях сохраняя свою жизнеспособность.


Часто в распространении возбудителей болезней растений участвуют насекомые (спорынья ржи), клещи (махровость смородины), нематоды (вирусная кольцевая мозаика малины). Распространение болезней растений человеком происходит при проведении ручных операций по уходу за растениями (при выламывании пасынков томата переносится вирус табачной мозаики), при механических обработках (с прилипшими на деталях культиватора частицами почвы по полю распространяются споры возбудителя рака картофеля). Кроме того, возбудители болезней растений могут быть завезены в новые районы и страны с семенным и посадочным материалом, с сельскохозяйственной продукцией и сырьем.
Вирусы, вызывающие заболевания, и методы защиты растений
Вирусы состоят из одиночной или двойной нити нуклеиновой кислоты (РНК, реже ДНК), окруженной белковой оболочкой—капсидом. Нуклеиновая кислота является носителем наследственных признаков патогена и определяет характер его инфекционное, а белок играет защитную роль и определяет способность вируса проникать в ткани растения-хозяина. Покоящиеся частицы вирусов — вирионы — разнообразны по форме (палочковидные, нитевидные, сферические, бацилловидные), имеют размеры от нескольких десятков до сотен нанометров (нм).
В настоящее время в мире принято англоязычное обозначение для наименования вирусов, часто сокращенное. Основу классификации вирусов, которые вызывают заболевания растений, представляют семейства и роды, которые упорядочены в основном в соответствии с видом нуклеиновой кислоты, характером генома, морфологией частиц, способом передачи и кругом растений-хозяев.


Например, типичный представитель рода (ранее группа) Tobamovirus из семейства Virgaviridae — вирус табачной мозаики (Tobacco mosaic virus), имеет размер вирионов 18×300 нм, состоящих из одноцепочечной РНК и белка с молекулярной массой (м. м.) 18 кДа (килодальтон), широкий спектр растений-хозяев, передается механически и семенами, представлен многочисленными штаммами.
Основное направление защиты растений от вирусных болезней — профилактические мероприятия. Размножение вирусов настолько тесно связано с обменом веществ клетки растения-хозяина, что непосредственное избирательное воздействие какими-либо препаратами на самого патогена отрицательно отражается и на растительной клетке. В связи с этим защитные мероприятия от вирусных заболеваний растений включают: организационно-хозяйственные методы; карантин; использование здорового семенного и посадочного материала (его получают с использованием термотерапии, химиотерапии и культуры апикальных меристем in vitro); селекционный метод по созданию устойчивых сортов и гибридов; борьбу с переносчиками и растениями-резерваторами; использование препаратов, повышающих иммунитет растений (иммуномодуляторов); преиммунизацию (искусственное заражение слабоагрессивными штаммами вируса) и некоторые другие меры. В маточных насаждениях в питомниках регулярно проводят фитосанитарные обследования с выбраковкой пораженных вирусами растений; соблюдают пространственную изоляцию поражаемых культур, маточных и товарных насаждений.
Симптомы и диагностика заболеваний растений, пораженных вирусами
Основными симптомами вирусных болезней — вирозов — являются угнетение роста, изменение окраски (чаще мозаики), деформация (морщинистость, скручивание, нитевидность, измельчение или нарушение зубчатости листовых пластинок, реже цветков и плодов), некрозы на листьях, стеблях, плодах, нарушение функций репродуктивных органов. Эти симптомы проявляются самостоятельно или в комплексе.


Мозаика плодов, лепестков цветов, а также хлоротичные кольца, дуги, извитые узоры являются достоверными симптомами многих вирусных болезней (шарка сливы, кольцевая мозаика томата). В некоторых случаях на растениях, зараженных вирусами, симптомы проявляются очень редко или не появляются вообще (вирус-Х картофеля), т. е. имеет место латентная, бессимптомная инфекция. Филогенетическая специализация вирусов может быть как узкой (вирус зеленой крапчатой мозаики огурца), так и широкой (вирус обыкновенной мозаики огурца).
К вирусам неприменимы традиционные микробиологические методы исследований. Основными методами их диагностики являются: визуальный (по внешним признакам); с помощью растений-индикаторов; метод электронной микроскопии; метод электрофореза; серологическая, или иммунологическая, диагностика, чаще иммуноферментный анализ (ИФА); молекулярно-генетические методы, чаще полимеразная цепная реакция (ПЦР). Для массовой диагностики вирусных инфекций растений в настоящее время используют модификации последних двух методов.
Размножение вирусов, которые вызывают заболевания растений
Механизм размножения вирусов — саморепликация, отличается от размножения других микроорганизмов. В клетках зараженного растения после отделения нуклеиновой кислоты вируса от его белковой оболочки происходит репродукция вируса путем синтеза отдельных молекул нуклеиновых кислот и белков, а затем последующая сборка из них вирионов. Вирусы влияют на генетический аппарат растения, изменяя его работу в сторону синтеза необходимых вирусу нуклеотидов и аминокислот. Некоторые вирусы размножаются в цитоплазме, а другие — в ядре клетки. Впоследствии происходит перемещение вирусных частиц из клетки в клетку по плазмодесмам, по межклеточной жидкости. Также вирусные частицы разносятся по всему растению с током питательных веществ по флоэме, особенно интенсивно при цветении и плодоношении; в меньшей степени они распространяются по тканям ксилемы. Иногда вирионы агрегатируют друг с другом, образуя в зараженных клетках вирусные включения (кристаллы Ивановского).
Вирусы, поражающие растения, не могут свободно проникнуть в растительную клетку, покрытую целлюлозной оболочкой, поэтому заражение растения возможно через мелкие ранки без грубых повреждений клеток при механической передаче и с помощью переносчиков через плазмодесмы от уже инфицированной соседней клетки. Распространение вирусов происходит при вегетативном размножении (все вирусы картофеля), семенами (вирус обыкновенной мозаики фасоли), контактно-механическим путем (вирус томатной мозаики), прививкой (шарка сливы), пыльцой (вирус кустистой карликовости малины), насекомыми (вирус мозаики свеклы), клещами (вирус полосатой мозаики пшеницы), почвенными нематодами, паразитирующими на корнях растений (вирус кольцевой мозаики томата), некоторыми грибами и псевдогрибами, в цикле которых образуются зооспоры (почвопередающийся вирус мозаики пшеницы), и растением-паразитом — повиликой (вирус мозаики люцерны). В агроценозы вирусы обычно попадают с семенным и посадочным материалом, а также с переносчиками. Большинство фитопатогенных вирусов распространяется насекомыми с колюще-сосущим ротовым аппаратом (тли, цикадки, белокрылки, трипсы, клопы), клещи также часто бывают переносчиками вирусов.
Основными источниками сохранения вирусов являются многолетние растения, в т. ч. сорняки. Они могут сохраняться в органах вегетативного размножения, семенах. Эта передача имеет важные экономические последствия, так как способствует сохранению вирусной инфекции длительное время и распространению на дальние расстояния. В переносчиках (насекомые, клещи, нематоды, покоящиеся споры грибов) вирусы также могут сохраняться в неблагоприятный период; единичные вирусы, стойкие к условиям внешней среды, — с растительными остатками и в почве, в гидропонных системах (вирусы мозаики томата).
Вироиды, поражающие растения
Вироиды — субмикроскопические неклеточные организмы, подобные вирусам, но лишенные капсида и состоящие только из низкомолекулярной одноцепочной нуклеиновой кислоты (РНК). Приоритет открытия вироидов принадлежит Теодору О. Динеру, который в 1971 году при изучении клубней картофеля, зараженных веретеновидностью, установил новый тип патогенов. Вироиды, как и вирусы, имеют английские названия. К настоящему времени известно более 30 вироидов, вызывающих такие заболевания, как веретеновидность (готика) клубней картофеля, экзокортис цитрусовых, карликовость хризантем, бледноплодность огурца, кустистость верхушки томата, карликовость хмеля, желтая крапчатость винограда. Основными симптомами вироидных болезней являются угнетение роста, изменение окраски (хлороз, антоцианоз), деформация различных органов.
Их распространение происходит с посадочным материалом, с семенами, контактно-механическим путем, а сохраняются они в многолетних растениях, органах вегетативного размножения и семенах. К основным методам диагностики вироидов относят визуальную диагностику, метод растений-индикаторов, электронную микроскопию, метод гель-электрофореза, ПЦР-анализ. Защита растений от вироидных болезней сходна с защитой их от вирусных патогенов. Основной упор должен быть сделан на профилактические мероприятия, такие, как использование здорового посадочного и семенного материала, отбор устойчивых клонов и сортов, дезинфекция режущих инструментов, используемых при уходе. Против вироидов практически неэффективны термотерапия и получение растений in vitro из верхушечных меристем.
Диагностика болезней растений и современные технологии

Обзор
Иллюстрация с обложки книги Ю.Т. Дьякова «О болезнях растений»
Автор
Редакторы
Статья на конкурс «био/мол/текст»: Фитопатология — это наука о болезнях растений, о том, как с этими недугами бороться и предупреждать их возникновение. Чтобы побеждать болезни, надо их правильно, точно и своевременно диагностировать. Есть много методов такой диагностики; в последние десятилетия, с развитием молекулярных методов анализа, активно разрабатываются всё новые и новые, постепенно вытесняя из практики классические подходы. О трудностях диагностики болезней растений и об основных путях их преодоления и будет рассказано в этой статье.

Конкурс «био/мол/текст»-2013
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2013 в номинации «Лучший обзор».
Спонсор конкурса — дальновидная компания Thermo Fisher Scientific. Спонсор приза зрительских симпатий — фирма Helicon.
Чем болеют растения?
Для начала несколько слов о том, от чего, собственно, специалистам приходится защищать сельскохозяйственные растения. Причинами заболевания растений могут быть как факторы среды (летняя засуха или зимние морозы, недостаток питательных веществ в почве или их избыток и т.п.), так и различные паразитические организмы (бактерии, вирусы, грибы, круглые черви (нематоды) и даже другие растения).
Грибы, бесспорно, являются основными патогенами культурных растений. Известно, например, что из 162 серьёзных заболеваний в Центральной Европе 135 (83%) вызываются грибами [2]. Фитопатогенные грибы — многочисленная группа; их описано свыше 10 000 видов, различных по систематическому положению, степени паразитизма, специализации и т.д. [3]. Они широко распространены в природе и при благоприятных для их развития условиях наносят значительный урон урожаю и сельскохозяйственным продуктам при хранении. Даже самые осторожные оценки говорят об уничтожении болезнями 10–20% потенциального урожая; без контрмер масштабы этих потерь резко возросли бы [2].
Именно о проблемах диагностики болезней растений, вызываемых фитопатогенными грибами, пойдёт речь в данной статье.
Врага надо знать в лицо
Зачем же нужно, с одной стороны — обнаружение, а с другой — быстрое и точное (желательно — до вида, или даже расы) определение фитопатогенных грибов?
На данный момент самым распространённым методом борьбы с фитопатогенными грибами является обработка растений фунгицидами. Понятно, что невозможно защитить культуры от всех возможных потенциальных угроз: это и сложно, и экономически невыгодно, да и для окружающей среды далеко не полезно. Именно поэтому важно знать, желательно — своевременно, с чем именно придётся бороться. Чем раньше обнаружена болезнь, тем больше шансов, что, приняв соответствующие меры, удастся её победить. Это верно для заболеваний как человека, так и растений. Кстати, точное определение вида грибов важно ещё и в довольно неожиданной области — реставрации деревянных строений — поскольку используемые там антисептические меры также очень сильно зависят от типа поражения [4].
Кроме этого, идентификация фитопатогенных грибов необходима для изучения их таксономии и эволюции, их взаимоотношений с растениями-хозяевами, генетических основ восприимчивости и устойчивости растений, что, в конечном счете, должно помочь в разработке способов борьбы с патогенами и в селекции растений, невосприимчивых к болезням [5].
И, наконец, крайне важна сертификация зерна и посадочного материала в рамках карантинных программ. Известно, что фитопатогенные грибы могут распространяться многими путями — как естественными (с током воздуха, водой, насекомыми, животными), так и при помощи человека, перевозящего заражённые растения или их части не только между различными странами, но и между континентами. Зачастую такое перемещение приводит к неожиданному и масштабному распространению заболеваний.
Например, пузырчатая ржавчина (Cronartium ribicola) была эндемична для Альп и востока России. Этот паразит, в цикле развития предполагающий обязательную смену хозяев, обитает круглый год на пятихвойных соснах, а летом поражает листья смородины; ни в одном из исходных ареалов он не причинял серьёзного ущерба. Однако веймутова сосна, завезённая в начале XVIII века из Америки в ряд областей Европы, оказалась крайне восприимчивым хозяином для данного гриба. За счёт этого распространившаяся инфекция причинила большой вред культурам смородины и высаженным веймутовым соснам, а в 1909 году была завезена с их рассадой в Америку, где встретила многочисленных хозяев для обеих фаз развития. Здесь стали страдать, прежде всего, лесообразующие пятихвойные сосны. Поэтому, чтобы разорвать инфекционную цепь паразита с обязательной сменой хозяев, пытаются уничтожать дикорастущие виды смородины [2].
Ещё один показательный пример: возбудитель голландской болезни вяза (Ophiostoma ulmi) уже в XX столетии был занесён из континентальной Европы в Северную Америку. Начиная примерно с 1970 г., после того, как он был завезён в Великобританию, он успел уничтожить половину английских вязовых насаждений [2]. Теперь этот вид встречается и в России.
Для того чтобы избежать подобного впредь, созданы списки карантинных организмов, и при перемещении растений или их семян между странами (или даже частями одной страны) обязательно проводится их обследование.
«Классические» методы диагностики и трудности в их применении
Как только что было показано, идентификация фитопатогенных грибов крайне важна, возник вопрос — каким образом она производится?
Наиболее простой способ — это идентификация патогена по внешним признакам заболевания (симптомам), то есть по тому воздействию, которое он оказывает на поражённое растение [6]. Но здесь проблема в том, что к одним и тем же повреждениям растения-хозяина могут приводить совершенно разные микроорганизмы, отличающиеся разной устойчивостью к фунгицидам, вредоносностью и другими характеристиками. Как пример, здесь можно привести три листовые пятнистости пшеницы (рис. 1).

Рисунок 1. Листовые пятнистости пшеницы. Слева — септориоз листьев пшеницы (возбудитель — Mycosphaerella graminicola). По центру — септориоз листьев и колоса пшеницы, проявление на листьях (возбудитель — Phaeosphaeria nodorum). Справа — жёлтая пятнистость пшеницы (возбудитель — Pyrenophora triticirepentis). Обратите внимание: несмотря на то, что это разные заболевания, поражения листьев очень похожи.
Ещё одна проблема заключается в том, что далеко не все заболевания проявляются сразу же после заражения растения. Например, возбудитель пыльной головни ячменя (Ustilago nuda) обычно проникает во время цветения пшеницы в формирующуюся зерновку. Гриб не препятствует формированию зародыша, само зерно развивается нормально, ничем внешне не отличаясь от здорового. Мицелий зимует в зерновке. Весной одновременно с прорастанием семян происходит и рост мицелия, который по мере роста растения распространяется по различным его органам. Проявляется заболевание только в период колошения. При этом разрушаются все части колоса, превращаясь в чёрную споровую массу, после распыления которой остаются лишь ости и стержень колоса (рис. 2) [8].

Рисунок 2. Пыльная головня ячменя: поражённое соцветие со спорами
Стандартный для фитопатологов подход при определении фитопатогенных грибов — это выделение их в чистую культуру на какой-либо питательной среде, получение характерных образований (чаще всего это, конечно, спороношения) и затем идентификация гриба под микроскопом.
Но здесь возникают определённые трудности. Основная из них заключается в том, что далеко не все паразитические грибы возможно культивировать на искусственных питательных средах: многим требуется наличие живых тканей растения-хозяина, либо присутствие других представителей сложного сообщества [10]. Но даже если гриб удаётся выделить в культуру, следующий вопрос — это то, сколько времени понадобится, чтобы добиться от него появления спороношения. Например, возбудитель белосоломенной болезни пшеницы и ржи (Gibellina cerealis), хотя и хорошо культивируется, даёт спороношение только после четырёх–пяти недель роста. Естественно, что меры по борьбе с патогеном необходимо принимать сразу после его обнаружения, а не через месяц, когда может оказаться, что спасать уже нечего.
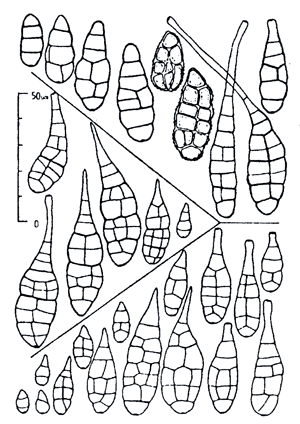
Рисунок 3. Сравнение конидий типовых образцов Alternaria longipes (вверху), Alternaria tenuissima (в центре), Alternaria alternata (внизу). Видно, что на основе сравнения только формы конидий этих трёх видов однозначно различить их крайне сложно. При идентификации видов в данном случае специалист использует не только форму конидий, но и другие признаки (например, способ образования конидий, их взаимное расположение и т.п.).
И даже с определением тех фитопатогенных грибов, спороношения которых получить сравнительно просто, могут возникать сложности. К примеру, идентификация многих микромицетов сопряжена с рядом трудностей, таких как сходство морфологических характеристик разных видов и одновременно внутривидовая вариабельность признаков. Несмотря на внешнее сходство, возбудители могут значительно отличаться по патогенности, токсигенности, степени специализации, генетике взаимоотношений с растением-хозяином, вредоносности, чувствительности к фунгицидам и т.д. То есть разные виды обладают совершенно разными экологическими особенностями и хозяйственной значимостью [12]. Хорошим примером здесь является определение различных видов рода Alternaria (рис. 3). Очевидно, что для идентификации до вида нужны достаточно широкие познания в данной области и немалый опыт работы с исследуемым фитопатогеном.
Ещё один способ, пригодный для обнаружения некоторых фитопатогенных грибов, заключается в смыве с субстрата, фильтрации и микроскопическом определении (и даже подсчёте, что даёт количественные данные) их спор. Чаще всего, таким способом оценивается количество грибных спор в зерне или в почве. Несмотря на то, что идентификация до вида на основании одних только спор чаще всего затруднена, этот способ широко применяется, а для анализа получаемых при помощи микроскопа изображений разрабатываются специальные компьютерные программы [14]. Например, таким образом определяют заражённость зерна возбудителем твёрдой головни (Tilletia caries) (рис. 4) [15]. Несмотря на использование компьютерных технологий, этот метод весьма трудоёмок и не подходит для исследования большого количества образцов.

Рисунок 4. Зерновки, поражённые твёрдой головнёй пшеницы
Молекулярная биология на службе фитопатолога
Во всех описанных случаях на помощь исследователям могут прийти широко развивающиеся в последнее время молекулярные методы анализа. Сейчас в основе большинства из них лежит применение ELISA (enzyme-linked immunosorbent assay, иммуноферментный анализ) [11], либо ПЦР (полимеразная цепная реакция, polymerase chain reaction) [17].
Иммуноферментный анализ состоит из двух основных этапов: иммунной и ферментативной реакций. Иммунная реакция заключается в специфическом связывании характерного для данного микроорганизма антигена с диагностическим антителом. Ферментативная реакция необходима для обнаружения этого связывания. Как правило, она сопровождается изменением цвета, причём степень этого изменения может быть использована для определения количества присутствующего антигена.
Есть много модификаций данного метода, бóльшая часть из которых применяется для обнаружения возбудителей болезней растений [16]. Особенно интересен так называемый «lateral flow assay» (анализ капли, растекающейся в радиальном направлении) (рис. 5), отличающийся высокой скоростью анализа и не требующий никакого специального оборудования или знаний. Используя такой набор (имеющий карманный формат), можно провести анализ непосредственно «в поле», так что не нужно даже отправлять образцы в лабораторию.
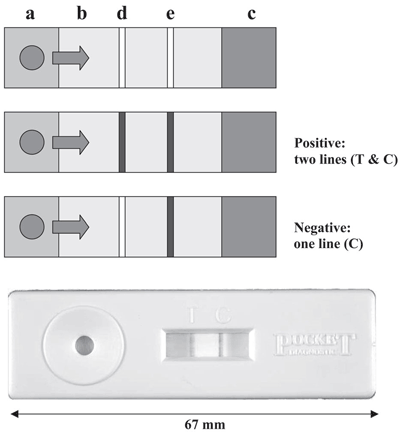
Рисунок 5. Прибор CSL Pocket Diagnostic TM lateral flow immunodiagnostic kit. Растительный экстракт помещается на площадку (a), которая содержит латексные шарики, покрытые специфическими антителами; смесь мигрирует вдоль мембраны (b) к абсорбирующей поверхности (c). При этом имеющиеся в растворе целевые антигены связываются со специфичными антителами на латексных шариках. Мембрана содержит полосу антител, отличающихся необходимой специфичностью (измерительную полосу) (d) и полосу других антител, которые связываются с первыми антителами (контрольную полосу) (e). Латексные шарики, содержащие связанный антиген, задерживаются в тестовой зоне, давая видимую линию, тогда как излишние латексные шарики, которые не содержат антигена, задерживаются в контрольной зоне, показывая, что анализ работает. Наличие двух линий соответствует положительному результату (positive), наличие только одной линии (контрольной) говорит о негативном результате (negative).
Основанные на иммуноферментном анализе методы широко применяются для обнаружения вирусов (в том числе поражающих растения) и значительно реже — для идентификации грибов и бактерий. Основной причиной этого является трудность получения антител с необходимой специфичностью: строение клеточных стенок грибов и бактерий гораздо сложнее, чем вирусного капсида, к тому же может изменяться в ходе их жизненного цикла. В результате получаемые антитела могут оказаться специфичны как сразу к большой группе видов, так и исключительно к отдельным жизненным формам данных микроорганизмов. Тем не менее, основанные на ELISA методы идентификации фитопатогенных грибов всё же разрабатываются: например, существует метод идентификации спор уже упоминавшейся в данной статье твёрдой головни [19].
ПЦР — это ферментативная реакция, в результате которой происходит накопление большого количества копий какого-либо не слишком большого (чаще всего, 200–1500 пар нуклеотидов) фрагмента ДНК. Так как ДНК любого организма содержит как вариабельные (отличающиеся даже у близкородственных организмов), так и консервативные (сходные у эволюционно далёких видов) участки, возможно на основе выбора диагностического участка варьировать специфичность протекающей реакции.
Таким образом, данный метод позволяет обнаруживать последовательности нуклеиновой кислоты, специфичные для конкретного организма или группы сходных организмов и, тем самым, выявлять его (их) присутствие в анализируемой пробе. Методы, основанные на ПЦР, позволяют идентифицировать патогенные виды как в чистой культуре, так и непосредственно в растительном материале, минуя этап изоляции грибов [20]. Как пример, здесь приведены результаты ПЦР, разработанной для идентификации грибов рода Pyrenophora (рис. 6), представители которого являются возбудителями жёлтой пятнистости злаков, в частности — пшеницы (рис. 1).
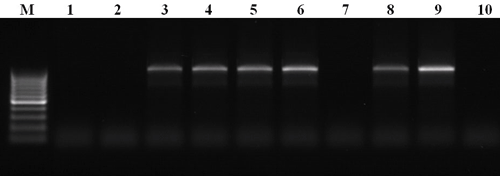
Рисунок 6. Разделённые при помощи электрофореза продукты ПЦР, разработанной для идентификации грибов рода Pyrenophora. М — маркер, представляющий собой набор фрагментов ДНК известного размера, 1–10 — ДНК, выделенная из различных образцов листьев пшеницы, поражённых листовыми пятнистостями. Здесь продукт реакции (фрагмент ДНК известного размера) должен наблюдаться только в том случае, если в образце присутствует ДНК целевого организма, а именно — гриба рода Pyrenophora. В итоге видно, что растения под номерами 3–6, 8 и 9 больны жёлтой пятнистостью, а остальные — каким-либо другим внешне схожим заболеванием.
Существует достаточно много модификаций метода ПЦР, большинство из которых применяется в изучении возбудителей болезней растений. Например, RAPD и RFLP анализы используются для уточнения родственных связей между различными грибами; ПЦР, специфичная для ДНК представителей отдельных родов или видов — для идентификации фитопатогенов (в том числе — в форматах nested и multiplex); ПЦР с регистрацией в режиме реального времени (real-time PCR) — для определения количества присутствующей целевой ДНК.
Рассмотрим подробнее один из самых перспективных методов на основе ПЦР — ПЦР с регистрацией в режиме реального времени (рис. 7). В отличие от большинства других форматов ПЦР, он позволяет не только констатировать факт присутствия ДНК целевого патогена, но и измерить её количество. В качестве примера здесь приведено определение в двух образцах количества ДНК ещё одного возбудителя листовой пятнистости.
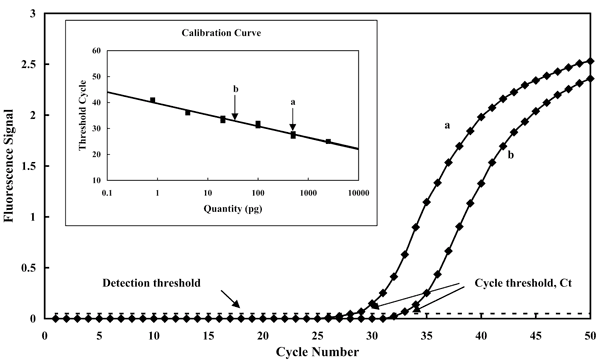
Рисунок 7. ПЦР с регистрацией в режиме реального времени. В ходе реакции непрерывно измеряется флуоресценция пробы (Fluorescence Signal), которая увеличивается по мере накопления продукта реакции (время реакции здесь выражено в числе циклов реакции — Cycle Number). Чем больше целевой ДНК было в смеси на момент начала реакции, тем быстрее происходит накопление продукта, и, соответственно, тем меньше будет номер цикла (cycle threshold, Ct), при котором флуоресценция пробы достигнет порогового значения (Detection threshold). Количество ДНК в исходной пробе определяют по калибровочному графику (приведён на вставке), соотносящему исходное количество целевой ДНК (Quantity) со значениями Ct. В данном случае на графике показано изменение во времени флуоресцентного сигнала для двух образцов, содержащих ДНК Mycosphaerella graminicola (рис. 1), причём кривая «а» соответствует образцу с большей концентрацией.
Интересно применение данного метода для анализа заражённости зерна твёрдой головнёй (рис. 4): при наличии соответствующих калибровочных графиков возможно получение результатов в виде числа спор, имеющихся в образце [7].
Ложка дёгтя в бочке мёда
Хотя преимущества и перспективы применения молекулярных методов идентификации сложно переоценить, на пути их практического использования имеется целый ряд трудностей. Несмотря на универсальность методов при конечном анализе, для их разработки и проверки требуется достаточно много времени и немалая экспериментальная база. Основной проблемой здесь является отсутствие возможности чисто теоретически оценить специфичность разрабатываемых методов.
Ещё одна сложность является прямым следствием высокой чувствительности данных методов (особенно — основанных на ПЦР). Ведь если анализ позволяет выявить в образце присутствие даже одной споры фитопатогенного гриба, или всего лишь нескольких копий его ДНК, то и для получения ложноположительного результата оказывается достаточно даже минимального загрязнения изначально «чистого» образца. Следовательно, требования к стерильности оказываются крайне высоки на всех стадиях анализа, но всё равно всегда приходится использовать отрицательные контроли.
Ну и самая большая проблема всех описанных в данной статье методов — это цена, ограничивающая их широкое применение в условиях небогатых российских хозяйств.
Несколько слов о будущем
Несмотря на все имеющиеся проблемы, молекулярные методы анализа интенсивно развиваются (о чём можно судить хотя бы по числу публикаций на соответствующие темы, которое с каждым годом становится всё больше). Старые методы постоянно совершенствуются, в то же время разрабатываются новые (например, метод биочипов [21] и секвенирование следующего поколения [22]), а цена одного анализа становится всё ниже. Поэтому можно надеяться, что не за горами то время, когда все упоминавшиеся в данной статье методики и их более совершенные аналоги действительно найдут широкое применение и облегчат жизнь фитопатологов и агрономов.
Источник https://www.pesticidy.ru/dictionary/The_causative_agent_of_the_disease
Источник https://kvetok.ru/vrediteli/infektsionny-e-i-virusny-e-bolezni-rastenij-osobennosti-patogeneza
Источник https://biomolecula.ru/articles/diagnostika-boleznei-rastenii-i-sovremennye-tekhnologii



