Устойчивость к болезням

У стойчивость растений к болезням, или иммунитет, может проявляться в абсолютной невосприимчивости либо в относительной устойчивости к заболеваниям. В первом случае возбудитель не влияет на состояние растения. Например, шишки сосны никогда не поражаются ржавчиной, а хвойные породы – мучнистой росой.
Врожденный и приобретенный
Причины устойчивости различны, и в зависимости от их характера выделяют врожденный и приобретенный иммунитет.
Врожденный иммунитет передается по наследству, но может изменяться под влиянием многих факторов (состояние растений, агрессивность патогена, условия внешней среды). Растения, обладающие врожденным иммунитетом, по-разному противостоят заражению, т. е. внедрению возбудителя. В одних случаях препятствием для проникновения инфекционного начала служат свойства растения, присущие только ему и существующие независимо от наличия или отсутствия патогена. Такой иммунитет называют пассивным. В других случаях способность противостоять заражению проявляется в быстрой реакции растения только в момент внедрения возбудителя. Такой иммунитет называют активным.
Степень пассивного иммунитета разных растений зависит от особенностей внешнего и внутреннего строения тканей, а также от их физиологических и биохимических свойств.
Физическая преграда
Толщина кутикулы (тонкая пленка, покрывающая эпидермис) имеет большое значение в устойчивости к болезням, возбудители которых проникают в ткани через кутикулярный слой. Чем он толще, тем большее препятствие представляет для проникновения инфекции. Например, барбарис Тунберга и б. удлиненный (продолговатый) более устойчивы к мучнистой росе и ржавчине, чем б. обыкновенный, у которого толщина кутикулы меньше. Менее восприимчивы к ржавчине чайно-гибридные, чайные и плетистые розы, у которых кутикула листьев толще, чем у других видов. Благодаря толстой кутикуле роза морщинистая обладает высокой устойчивостью (близкой к абсолютной) к мучнистой росе. Этим же фактором обусловлена и возрастная устойчивость лиственных пород к данной болезни.
С возрастом толщина кутикулы увеличивается и соответственно снижается восприимчивость листьев и побегов к заболеванию. Так, листья и побеги прикорневой поросли в сильной степени поражаются мучнистой росой и усыхают, тогда как материнское растение практически не страдает от болезни.
Густое опушение и восковой налет на поражаемых органах не только служит физической преградой для проникновения инфекции, но и препятствует попаданию сохранению на них влаги, необходимой для заражения. Так, хвоя голубых форм ели, покрытая восковым налетом, более устойчива к ржавчине и шютте. Этим же объясняется меньшая восприимчивость лиственницы японской к шютте (мериозу).
Ряд возбудителей проникает в ткани растения через устьица и чечевички, являющиеся естественными воротами инфекции. Небольшое количество и маленькие размеры этих отверстий снижают вероятность проникновения возбудителя в ткани.
Нередко устойчивость растений к болезням может зависеть от толщины клеточных стенок. Так, многолетними исследованиями установлено, что наименее восприимчивы к корневой губке (Heterobasidion annosum) виды сосны, отличающиеся утолщенными клеточными стенками в древесине.
На устойчивость древесных растений к болезням может оказывать влияние габитус кроны. В раскидистых ажурных кронах, которые хорошо освещаются и проветриваются, создается более сухой микроклимат, неблагоприятный для развития возбудителей. Например, формы тополя с пирамидальной плотной кроной сильнее поражаются бурым цитоспоровым (Cytospora chrysosperma) и дискоспориевым (Discosporium populeum) некрозами.
Степень пассивного иммунитета разных растений зависит от особенностей внешнего и внутреннего строения тканей, а также от их физиологических и биохимических свойств.
Внутреннее содержание
Огромную роль в устойчивости растений играет состав клеточного сока и наличие в растительных тканях химических соединений, препятствующих проникновению патогенов или подавляющих их развитие (фенолы, алкалоиды, эфирные масла, смолы, дубильные вещества и др.). Так, установлено, что клеточный сок устойчивых к голландской болезни (Ophiostoma ulmi) видов ильмовых пород содержат вещества, в значительной мере подавляющие развитие возбудителя.
Немаловажное значение в изменении устойчивости древесных пород имеет содержание воды в тканях. Восприимчивость многих растений повышается при уменьшении влажности коры и древесины. Этим объясняется снижение устойчивости деревьев к сосудистым и некрозно-раковым болезням.
Из химических веществ следует отметить фенольные соединения, наличие которых повышает устойчивость сосны и ели к корневой губке.
Повышение устойчивости у древесных растений нередко обусловливается фитонцидами (биологически активные вещества, которые являются постоянным признаком растения, подавляющие рост и развитие патогенов). Например, менее восприимчивые к сосудистому микозу (Ophiostoma roboris) разновидности дуба характеризуются наиболее высокой фитонцидной активностью.
Барбарис Тумберга более
устойчив к мучнистой росе,
чем барбарис обыкновенный Роза морщинистая обладает высокой устойчивостью к мучнистой росе Восковой налет на хвое делает лиственницу устойчивой к шютте
Защитные реакции
Активные реакции растений на внедрение возбудителей проявляются в образовании защитных некрозов, возникновении антитоксинов и антиферментов.
Защитные некрозы представляют собой отмершие участки тканей, в которых патогены с высокой паразитической активностью развиваться не могут и погибают. Такая реакция проявляется при поражении листьев ржавчиной и мучнистой росой.
Антитоксические и антиферментные реакции выражаются в образовании фитоалексинов, активизации окислительных ферментов, образовании механических барьеров для проникновения патогенов. Фитоалексины – это вещества, которые образуются в тканях растения только при проникновении патогена и способны задерживать или полностью подавлять его развитие.
У многих растений, устойчивых к той или иной болезни, вырабатываются восстановительные ферменты, которые не только снижают активность выделяемых патогеном ферментов или полностью подавляют их образование, но и способствуют синтезу веществ, необходимых для восстановления клеток и тканей, разрушенных возбудителем.
Снижение восприимчивости древесных растений к болезням может быть связано с образованием механических барьеров, препятствующих распространению возбудителя, локализующих его. Например, реакция деревьев лиственных пород на внедрение опенка осеннего (Armillaria mellea) проявляется в образовании пробковой ткани на границе между здоровой и разрушаемой грибом ткани.
На устойчивость древесных растений к болезням может оказывать влияние габитус кроны.
Важное приобретение
Очень большое значение в повышении устойчивости древесных растений к болезням имеет приобретенный иммунитет. Он возникает после перенесенной болезни, а чаще – вследствие целенаправленного воздействия внешних факторов или химических (удобрения, микроэлементы) и биологических (вакцины) веществ.
Иммунитет, возникающий после перенесенной болезни, называется инфекционным и встречается крайне редко. Наиболее важная роль принадлежит неинфекционному иммунитету, который создается человеком посредством разных приемов, таких как агротехнические мероприятия, направленные на организацию оптимальных условий для развития растений, применение химических (химическая иммунизация) и биологических (биологическая иммунизация) веществ. Химическая иммунизация растений проводится чаще путем применения удобрений и микроэлементов, реже – антиметаболитов.
Наиболее важными для растений являются азотные, калийные и фосфорные удобрения, поскольку они играют большую роль в обмене веществ и действии физиологических функций. Внесение их в почву в оптимальном соотношении значительно повышает устойчивость растений к болезням, в том числе к инфекционному полеганию всходов, мучнистой росе, ржавчине, парше и др.
Важное значение в снижении восприимчивости растений имеют микроэлементы: бор, марганец, медь, цинк, железо, кобальт и др. Они способствуют утолщению покровных тканей и клеточных стенок, подавляют деятельность ферментов и токсинов, выделяемых патогеном, рост грибницы возбудителей болезней. Так, применение микроэлементов снижает восприимчивость древесных растений к полеганию, шютте сосны и лиственницы.
Из антиметаболитов (органические вещества, близкие по химической структуре к продуктам обмена веществ растений) применяют аргинин, анденозин, пиритиамин, гидрохинон и паранитрофенол. Известно, что два последних повышают устойчивость ильмовых пород к голландской болезни. Биологическая иммунизация проводится посредством введения в ткани растения вакцин (препараты из живых или убитых микроорганизмов и продуктов их жизнедеятельности). В результате в тканях растения образуются вещества, препятствующие развитию патогена или нейтрализующие выделяемые им токсины.
Применение микроэлементов снижает восприимчивость древесных растений к полеганию, шютте сосны и лиственницы.
К дискоспориевому некрозу наиболее устойчив тополь серебристый Хвоя голубых елей покрытая восковым налетом более устойчива к ржавчине и щюте Сосна кедровая сибирская очень восприимчива к пузырчатой ржавчине
Подбирая ассортимент
На деревьях и кустарниках развивается большое количество инфекционных болезней, но для каждой конкретной породы наиболее опасными являются отдельные виды или группы болезней, что следует учитывать при подборе ассортимента растений для посадки.
Для разных лиственных пород деревьев и кустарников, особенно для последних, опасными являются ржавчина, мучнистая роса и некоторые пятнистости листьев. Наиболее сильно ржавчиной поражается тополь белый, барбарис, роза. Высокой восприимчивостью к мучнистой росе отличаются дуб, клен, ясень, барбарис, жимолость, карагана древовидная (желтая акация), роза, сирень. Следует отметить, что абсолютно устойчива к болезни липа. К опасным и широко распространенным пятнистостям листьев относятся красно-коричневая боярышника (Phyllosticta michailowskoensis), бурая дуба (Discula umbrinella), коричневая каштана конского (Phyllosticta sphaeropsoidea), черная клена (Rhytisma acerinum), кремовая липы (Gloeosporium tiliae), черная розы (Marssonina rosae).
Наибольшую опасность для городских насаждений представляют сосудистые и некрозно-раковые болезни стволов и ветвей, приводящие к ослаблению и усыханию деревьев и кустарников. Массовое и быстрое усыхание ильмовых пород вызывает голландская болезнь, менее восприимчивым к которой считается вяз мелколистный.
Довольно часто причиной неблагополучного состояния тополя является дискоспориевый некроз (Discosporium populeum). Наиболее устойчивы к болезни разные формы белого тополя. Быстрое усыхание ивы вызывает диплодиновый некроз (Diplodina microsperma), поражающий преимущественно плакучие формы разных видов. К значительной потере декоративности и постепенному усыханию липы приводит поражение тиростромозом (Thyrostroma compactum).
Очень опасным заболеванием сосны является пузырчатая ржавчина, или ржавчинный рак (Cronarcium ribicola). Наиболее восприимчивы к ней сосна веймутова и с. кедровая сибирская. Значительный вред разным видам сосны в городских насаждениях в последние годы причиняет сферопсисовый некроз (Sphaeropsis sapinea). Наиболее высокая устойчивость к болезни отмечена у с. Банкса и с. скрученной.
Большое значение в повышении устойчивости насаждений к болезням имеет правильный подбор пород для конкретных почвенно-климатических условий. Несоответствие условий произрастания требованиям древесных пород приводит к снижению их устойчивости ко многим некрозно-раковым болезням, в том числе к цитоспорозу (Cytospora) и туберкуляриевому некрозу (Tubercularia vulgaris) лиственных пород, побеговому раку сосны (Scleroderris lagerbergii ). При этом следует отдавать предпочтение более устойчивым к тем или иным болезням видам, формам, сортам деревьев и кустарников.
Защитные некрозы – это отмершие участки тканей, в которых патогены развиваться не могут и погибают.
Делаем выводы
Таким образом, иммунитет растений очень сложное и многообразное явление, которое определяется особенностями растения-хозяина, возбудителя болезни и условиями внешней среды. Каждый из этих факторов обладает большим разнообразием и изменчивостью свойств на фоне особенностей внешних условий. Поэтому степень устойчивости растений к болезням не является постоянной и может повышаться или снижаться по целому ряду причин. К ним относятся: естественная и искусственная гибридизация (скрещивание) среди растений-хозяев, изменение агрессивности патогена (способности вызывать массовое поражение), условия внешней среды (высокие и низкие температуры, избыток и недостаток влаги в почве, дисбаланс питательных веществ, загрязнение окружающей среды, рекреационная нагрузка).
При создании насаждений необходимо проводить комплекс мероприятий, направленных на повышение их устойчивости к болезням. При этом большое значение имеет отбор пород, видов, разновидностей, сортов, устойчивых к опасным и широко распространенным заболеваниям.
Во избежание развития очагов болезней в насаждениях необходимо использовать для посадки растения без признаков поражения болезнями и повреждений различного характера. При посадке следует избегать загиба, повреждения корней и заглубления корневой шейки, что приводит к сильному ослаблению растений и снижению их устойчивости к болезням.
Очень важное значение в повышении устойчивости древесных растений к болезням имеет химическая иммунизация с применением удобрений и микроэлементов. Повышению устойчивости городских насаждений к болезням в значительной мере способствует снижение рекреационных нагрузок. В зонах с интенсивной рекреацией происходит сильное уплотнение почвы, обнажаются корневые лапы деревьев, увеличивается вероятность механических повреждений корней, стволов и ветвей, которые служат воротами инфекции. Это приводит к ослаблению деревьев и кустарников, снижению их устойчивости к корневым гнилям и некрозно-раковым болезням.
Массовое поражение листьев ржавчиной, мучнистой росой, пятнистостями, паршой приводит к ослаблению растений и снижению их устойчивости к некрозным болезням. Поэтому при систематически повторяющемся поражении листьев необходимо проводить опрыскивание растений в период вегетации фунгицидами.
Устойчивость растений: от молекул и генов к организму
Обзор
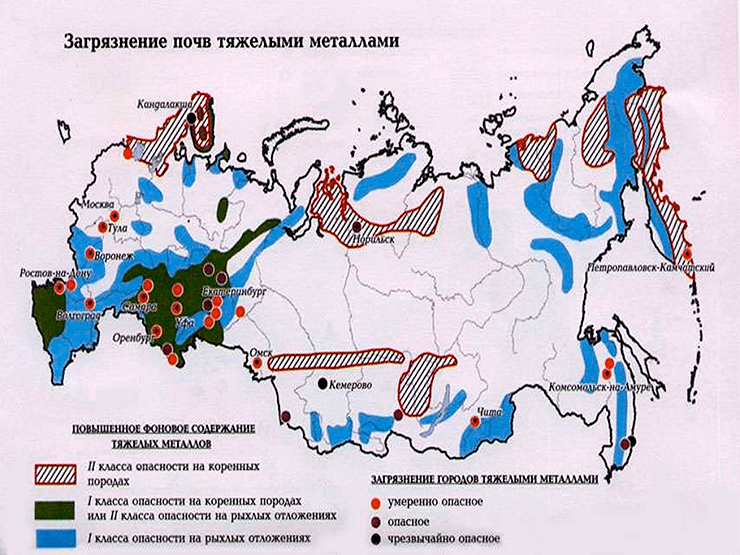
Карта загрязнения почв России тяжелыми металлами.
Автор
Редакторы
Статья на конкурс «био/мол/текст»: «Ухудшение экологической ситуации» — одна из тех проблем техногенного мира, которая ныне активно муссируется в СМИ и общественных дискуссиях. Частный случай этой проблемы — загрязнение окружающей среды тяжелыми металлами. Подчеркивается ядовитость этих веществ, угроза употребления загрязненной продукции для животных и человека. В связи с этим возникает вопрос: если эта проблема настолько серьезна, то почему же планета еще не превратилась в мертвую пустыню, а человечество не столкнулось с проблемой голода? Причина, по которой мы не наблюдаем столь серьезных последствий техногенеза, — механизмы, обеспечивающие устойчивость растений. Вопросу формирования этой устойчивости и способам ее исследования и посвящена эта статья.

«Био/мол/текст»-2016
Эта работа опубликована в номинации «Своя работа» конкурса «био/мол/текст»-2016.

Спонсор номинации — Future Biotech, проект, объединяющий профессионалов и энтузиастов в области биологии и биотехнологий.
Генеральным спонсором конкурса, согласно нашему краудфандингу, стал предприниматель Константин Синюшин, за что ему огромный человеческий респект!
Спонсором приза зрительских симпатий выступила фирма «Атлас».
Что такое металлы?
Металлы — это химические элементы, обладающие металлической связью и металлическим типом кристаллической решетки, в узлах которой находятся положительно заряженные ионы (атомы металла, потерявшие валентные электроны), а между ними расположен «электронный газ» — свободные электроны валентных энергетических уровней (s, p внешнего и d предвнешнего энергетических подуровней, а у актиноидов и лантаноидов — еще и f предпредвнешнего), которые и обеспечивают связь в кристалле, не принадлежа ни одному конкретному атому. К металлам относится большинство элементов периодической системы Д.И. Менделеева (95 из 118 известных на настоящий момент). Уникальное строение кристаллической решетки придает металлам ряд интересных свойств, таких как высокая электропроводность, ковкость, прочность, износоустойчивость и другие, что делает их незаменимым сырьем для многих отраслей народного хозяйства [1]. Как «наивно» определял эти элементы еще М.В. Ломоносов, «металлы — это твердые тела, которые ковать можно». Металлы используются с глубокой древности, и по мере развития общества диапазон известных и применяемых человеком представителей этого класса химических элементов только расширялся. И со всей уверенностью можно заявить, что человеческая цивилизация в ее нынешнем виде была бы невозможна, если бы не существовало металлов и их месторождений.
При этом наиболее интересным и востребованным в народном хозяйстве «классом» металлов являются так называемые тяжелые металлы (ТМ) — то есть такие, плотность которых превышает 5 г/см 3 и к которым относится большинство металлов. Это объясняется тем, что легкие металлы, являясь типичными представителями этого вида химических элементов, имеют и существенные недостатки. Благодаря тому, что у них существует только металлический тип связи, а валентные электроны внешних энергетических уровней могут легко отрываться в ходе химических реакций, они обладают высокой химической активностью и низкими механическими свойствами. Так, натрий по прочности и пластичности напоминает пластилин, быстро окисляется на воздухе, а в воде загорается. Все эти особенности делают его малопригодным для использования в чистом виде. Напротив, тяжелые металлы в большинстве своем лишены этих недостатков. Имея множество свободных d-орбиталей, они способны к образованию донорно-акцепторных связей по ковалентному типу, что обеспечивает им куда лучшие механические свойства. Также их валентные электроны прочнее удерживаются атомами, что снижает реакционную способность. Это открывает этим металлам дорогу к широкому применению в народном хозяйстве.
Чем опасны тяжелые металлы?
Однако у этих замечательных свойств со временем обнаружилась и обратная сторона. А именно то, что тяжелые металлы и их соединения оказались довольно токсичными для живых существ. Причин этому можно выделить несколько. Во-первых, это упомянутая выше возможность образования ковалентных связей. Ионы металлов способны связываться с биологическими молекулами, обладающими отрицательным зарядом — например, с карбоксильными остатками аминокислот. Во-вторых, известно, что активные центры ферментов нередко содержат ионы металлов. Металлы, имеющие химическое подобие с таким кофактором — например, как кадмий с цинком, — могут замещать нужный ион в активном центре, что ведет к нарушению функционирования фермента. Примером такого фермента может служить супероксиддисмутаза (SOD), содержащая (в зависимости от формы) в активном центре Zn, Mn, Fe. Эти элементы могут быть замещены на Cd, Cr и Ni соответственно [2]. В-третьих, тяжелые металлы способны вызывать «высаливание» белков, то есть выпадение в осадок растворимых белков за счет нарушения их нативной структуры. В-четвертых, известно, что ТМ способствуют повышенной выработке в клетке активных форм кислорода (АФК) [3], [4]. АФК реагируют с различными веществами, слагающими клетку, окисляя их (это явление называется окислительным стрессом), и способны привести даже к гибели организма. Поэтому, учитывая, что тяжелые металлы широко используются в народном хозяйстве, становится весьма актуальным вопрос изучения их воздействия на живые организмы и поиск путей защиты от этого токсического влияния. В качестве примера на рисунке 1а приведем схему, иллюстрирующую пути поступления в окружающую среду свинца — одного из тяжелых металлов. А на рисунке 1б — воздействие свинца на различные жизненные процессы растений [5].
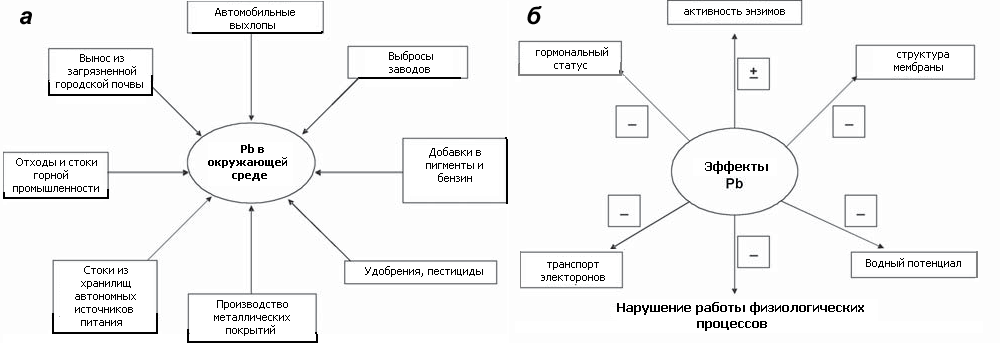
Рисунок 1. Поступление свинца в окружающую среду и его влияние на растения. а — Источники загрязнения окружающей среды свинцом. б — Общее представление о токсических эффектах свинца, оказываемых им на растения (за исключением водного потенциала, схема применима и к животным). Знаки «+» и «−» показывают, каков характер влияния свинца на конкретный процесс — позитивный или негативный.
рисунок автора статьи, на основе данных из [5]
В результате процессов, указанных на рисунке, в окружающую среду поступает большое количество ТМ. Так, с выхлопными газами в атмосферу поступает более 250 тыс. тонн свинца в год. Его содержание в дожде и снеге составляет до 300 мг/дм 3 . Ежегодно житель города поглощает около 45 мкг свинца, и его содержание в крови современного человека превышает таковое у первобытного в 100 раз [6]. Для человека один из возможных путей токсического действия свинца — замещение кальция в костях.
Из атмосферы ТМ вместе с осадками попадают в почву (в том числе сельскохозяйственных угодий), а затем и в продукты питания, произведенные на загрязненных территориях. Ареалы техногенных выбросов вокруг промышленных предприятий охватывают площадь 18 млн га, что соответствует 1% территории Российской Федерации. Площадь загрязненного ТМ почвенного покрова составляет 3,6 млн га (см. заглавный рисунок). Более миллиона гектаров сельскохозяйственных угодий загрязнено особо токсичными элементами (I класс опасности — Hg, Se, As и др.) и 2,3 млн га — токсичными (II класс опасности — Zn, Fe, Cu и др.) [7].
Что такое устойчивость к металлам?
Впрочем, как следует из эволюционной теории, наличие в среде обитания некоего стрессового фактора приводит к тому, что организмы, населяющие эту территорию, подвергаются давлению отбора, ведущего к ослаблению действия этого фактора [8]. Здесь мы подходим к такому феномену, как устойчивость к действию ТМ. Особенно это актуально для растений: если животные часто мигрируют и могут покинуть загрязненную территорию, то растения ведут прикрепленный образ жизни. При этом они непосредственно контактируют с загрязненной средой (почвой). Соответственно, со временем загрязненные территории должны были заселяться видами и линиями растений, устойчивых к воздействию стрессоров. Существуют две стратегии, позволяющие растениям успешно развиваться на загрязненной территории. Первая — «избегание» — развитие механизмов, направленных на недопущение попадания вредных соединений во внутреннюю среду растения. Вторая — «устойчивость» — развитие специфических механизмов, позволяющих растению успешно функционировать в присутствии этих веществ во внутренней среде организма [9]. Стоит заметить, что устойчивость является своего рода «палкой о двух концах»: она обеспечивается ценой существенного напряжения сил организма и порою снижением ряда важных жизненных показателей, поэтому на «чистых» территориях такие растения зачастую не выдерживают конкуренции с представителями обычных, неустойчивых линий [10]. Но на загрязненных ТМ территориях можно наблюдать такую картину: превращенные фактически в пустыню, они постепенно колонизируются устойчивыми расами разных видов растений (рис. 2).

Рисунок 2. Последствия загрязнения природных ландшафтов тяжелыми металлами. а — Вид района разработок медной руды в Копперхилле (США, штат Теннеси). Растительность практически отсутствует, в то время как раньше здесь произрастали леса. б — Вид долины Тайдслоу Рейк (Великобритания, графство Дербишир), мéста, где 200 лет назад добывали свинец; ныне она колонизирована устойчивыми к металлам расами широкого диапазона растительных видов.
Однако оставим вопрос устойчивости к ТМ дикорастущих видов растений и взглянем на проблему с иной стороны, более интересной с хозяйственной точки зрения: обратимся к теме устойчивости к ТМ сельскохозяйственных растений. Этот вопрос весьма актуален для нашей страны, поскольку проблема загрязнения агросферы ТМ, как мы уже убедились, существует (рис. 1б), и становится особенно значимым вопрос получения достаточного количества качественной и безопасной сельскохозяйственной продукции. Теме устойчивости сельскохозяйственных растений к действию ТМ посвящено довольно много работ, рассмотренных хотя бы в обзоре [2], однако здесь по-прежнему существует немало нерешенных проблем, и механизмы устойчивости растений всё еще нуждаются в дополнительном исследовании. Впрочем, чтобы не оставлять читателя в неведении, приведем схему ответа растительного организма на действие ТМ на примере свинца (рис. 3). При этом мы позволим себе здесь не касаться животных. Во-первых, потому, что эти механизмы в значительной степени будут дублироваться. Во-вторых, потому, что растения — основа любого биоценоза, и ТМ попадают в пищевую цепь сначала через организм продуцентов (т.е. растений), а уж потом переходят из них к животным. Поэтому можно считать, что если нам удалось разобраться с вопросами устойчивости растений, то мы в значительной степени уяснили для себя пути устойчивости экосистем в целом.
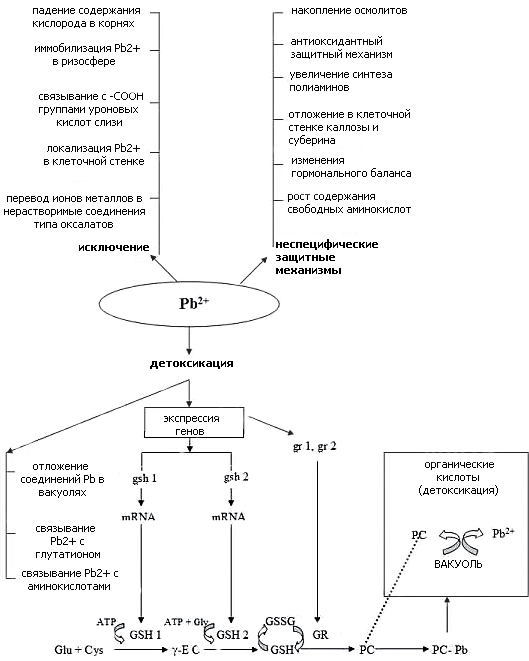
Рисунок 3. Схема ответа растительной клетки на токсическое действие свинца. Описание приведено в тексте.
рисунок автора статьи, на основе данных из [5]
Распознавание стрессора, то есть рецепция сигнала, приводит к активации пути передачи сигнала. Сигнал поступает в генόм, индуцируя или подавляя синтез тех или иных белков. Связанные с экспрессией генов ответные реакции клеток на действие стрессора интегрируются в ответ растения, выражающийся в ингибировании роста и развития растения и одновременно в повышении его устойчивости к действию стрессора. Способность растений удалять нежелательные ионы связана с падением содержания кислорода в корнях, эффективностью иммобилизации металлов в ризосфере, локализацией в клеточной стенке, связыванием с —СООН группами уроновых кислот слизи и отложением в виде оксалатов. Механизмы же детоксикации включают изоляцию свинца в вакуолях путем образования комплексов, связывания свинца глутатионом (GSH), аминокислотами (AA), фитохелатинами (PC). Синтез PC происходит совместно с сокращением количества глутатиона в клетках и ростом активности глутамилцистеинсинтетазы (GSH 1), глутатионсинтетазы (GSH 2) и глутатионредуктазы (GR). Повышенная активность GSH 1, GSH 2 и GR коррелирует с усилением экспрессии соответствующих генов (на рисунке 3 обозначенных как gsh 1, gsh 2, gr 1 и gr 2). Неспецифические защитные механизмы выражаются в том, что наличие свинца приводит к накоплению осмолитов, антиоксидантов, каллозы, суберина, аминокислот и изменению гормонального баланса.
Что такое генетические маркеры?
Прежде всего раскроем смысл самого термина «маркер». Под маркером понимают некий признак биологического объекта, который можно относительно легко выявить, и отражающий определенные внутренние особенности организма. В принципе, в качестве маркеров можно использовать и некоторые внешние особенности организма — как наиболее доступные для наблюдения: форму органа, окраску, опушенность и т.д., — что и делалось ранее [13]. Однако набор таких признаков ограничен по сравнению с числом генов, и сами они сильно зависят от многих условий и контролируются работой разных генов. Наиболее перспективны в этом плане биологические молекулы, обладающие полиморфизмом, то есть существующие в организме в виде нескольких изомеров. Каждый белок кодируется определенной нуклеотидной последовательностью, и наличие в ДНК тех или иных различий внутри одного биологического вида ведет к тому, что у отдельных индивидов этот белок существует в виде ряда изомеров, позволяющих судить о генетических особенностях организма. Поэтому в настоящее время используются молекулярные маркеры — белковые (изозимы, антигены, запасные белки) или ДНК. Здесь мы более подробно остановимся на первых.
В нашей работе [14] мы использовали изозимные маркеры. Предпочтение им было отдано потому, что ДНК-маркерам, несмотря на все их несомненные достоинства, свойственны следующие недостатки: дороговизна и трудоемкость методов анализа; высокая токсичность ряда часто используемых реагентов; необходимость строго следить за сохранением чистоты образца, в который ни в коем случае не должна попадать чужеродная ДНК; некоторые отмечаемые по результатам анализа генетические варианты могут не находить фенотипического проявления, что исключается в случае работы с белковыми маркерами, где наблюдается активность только реально работающих генетических систем.
У белковых маркеров есть и другие достоинства. Во-первых, белки — это первичные продукты генетических систем, поэтому путь от гена до белка значительно короче, чем до морфологических и даже обычных биохимических и физиологических признаков. По этой причине белковые маркеры подвержены значительно меньшей фенотипической изменчивости. Во-вторых, каждый белок представляет собой уникальный продукт соответствующей ему генетической системы и наилучшим образом отражает специфику этой системы. В-третьих, белок — обязательный продукт практически всех генетических систем [15]. Белки имеют ряд особенностей — от биологических до физико-химических, вроде заряда, конформации, диффузионной подвижности и других. Это позволяет использовать их в качестве маркеров не только генов, но и идентификации вида, сорта, линии живых организмов [16]. Так, в селекционной работе в качестве маркеров используют различные изомеры так называемых запасных белков, которые могут быть уникальными для отдельного сорта растения [17]. Белки играют важную роль в биологических процессах, через них генотип непосредственно трансформируется в фенотип. Как уже говорилось, белки обладают множественностью форм, которые отражают полиморфизм (множественность форм) биологического вида, поэтому исследователь может судить о генетических особенностях организмов и произвести их классификацию на основе белковых маркеров. Схема происхождения множественности форм белков показана на рисунке 4.
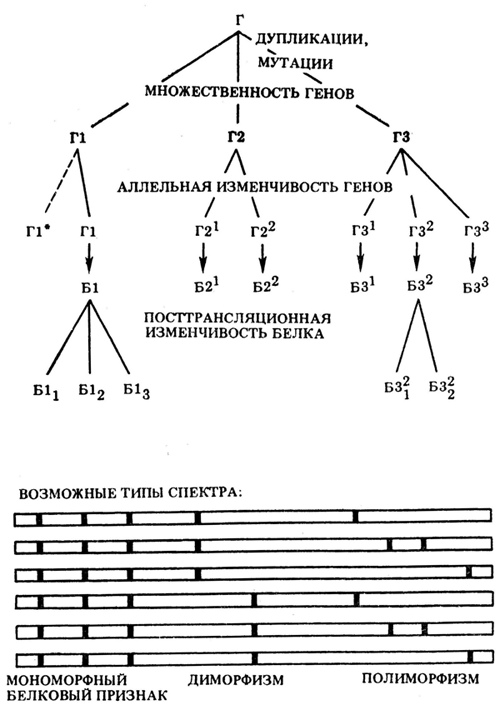
Рисунок 4. Происхождение множественности и полиморфизма белков. Б — белки, Г — гены, Г1* — выпавший или очень редкий аллель. Частота встречаемости типов спектра связана с частотой встречаемости аллелей и характером сцепления между генами.
Удобными объектами для исследования устойчивости считают ферменты, связанные с ответом живого организма на действие неблагоприятных факторов среды. Их преимущества перед иными возможными объектами состоят в том, что они являются продуктами реально работающих генов (в отличие от ДНК, которая может нести «молчащую» информацию, не находящую проявления в фенотипе) и напрямую связаны с реакцией на стресс (в отличие от запасных белков). Если у таких энзимов удастся обнаружить аллели, с большей вероятностью встречающиеся у устойчивых сортов, то можно полагать, что именно они и обуславливают эту устойчивость, лучше справляясь с негативным агентом. Примером такой работы может быть статья [18], посвященная исследованию изоформ супероксиддисмутазы, встречающихся у растений в зоне Восточно-Уральского радиоактивного следа (ВУРС).
Одним из подходящих объектов для такого исследования является яровой двурядный ячмень (Hordeum vulgare L.). Его выбор можно объяснить тем, что ячмень — сельскохозяйственная культура, известная с глубокой древности и возделываемая практически во всему миру, в самых разных географических, почвенных и климатических условиях. При этом она достаточно хорошо изучена: существуют исследования, касающиеся выхода цитогенетических нарушений [19], антиоксидантной активности [20], апоптоза [21] и физиологических процессов [22] у ячменя.
Исследование устойчивости сельскохозяйственных растений к ТМ
Автор этой статьи работает во Всероссийском научно-исследовательском институте радиологии и агроэкологии, в лаборатории радиобиологии и экотоксикологии растений. Интерес автора к рассматриваемой теме обусловлен тем, что она составляет тематику его исследования — изучение устойчивости ячменя к действию свинца [14], [23]. А выбор такой темы обусловлен тем, что она представляет существенный интерес как в практическом плане (создание устойчивых сортов), так и в теоретическом (изучение фундаментальных механизмов ответа живого организма на действие стресса). Общая схема этого исследования дана на рисунке 5. Как видно, она состоит из трех блоков: исследование ответа ячменя на действие диапазона концентраций свинца (на примере нескольких сортов); изучение ответа большого числа сортов на действие избранной концентрации Pb 2+ ; исследование причин формирования полиморфизма по устойчивости.
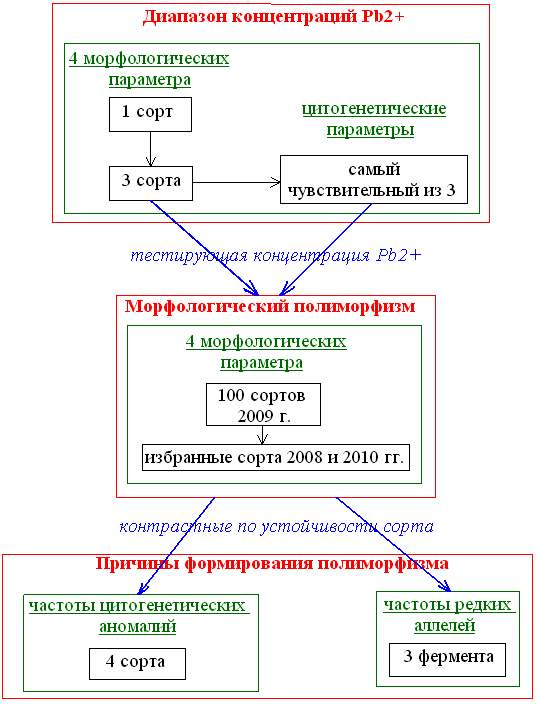
Рисунок 5. Схема экспериментов по исследованию влияния свинца на яровой ячмень
В первой части работы были проведены эксперименты по определению тестирующей концентрации свинца — дозы ТМ, которая уже вызывает значимое по сравнению с контрольным вариантом угнетение жизненных процессов растений ячменя, однако всё еще оставляет им возможность для развития. При такой концентрации ТМ в среде растение способно пройти все фенологические фазы развития и дать урожай, хотя и будет демонстрировать существенную степень угнетения. Этот показатель позволяет разделить сорта по устойчивости к поллютанту.
Определение тестирующей дозы ТМ проводили на основе анализа морфологических и цитогенетических показателей, чтобы сделать отбор комплексным, основанным не только на внешне видимых признаках, но и на более тонких особенностях растения. Поэтому первой частью исследования стало испытание трех сортов в широком диапазоне доз соли свинца. На основе анализа этих показателей мы оценили критическую концентрацию. Для уточнения дозы выполнили цитогенетическое исследование клеток апикальной меристемы корешков проростков наиболее чувствительного по результатам предшествующего эксперимента сорта.
На следующем этапе исследовали полиморфизм 100 сортов ячменя по морфологическим показателям при действии избранной дозы загрязнителя. Целью этапа было выделение групп наиболее устойчивых и чувствительных сортов. Для этого семена 100 сортов урожая 2009 г. проращивали в двух вариантах — на дистиллированной воде и на растворе, содержащем тестирующую концентрацию свинца. Итогом этого этапа работы стало выявление нескольких устойчивых и чувствительных сортов, особенности которых воспроизводились у урожаев трех лет.
Теперь возникает вопрос: какие тонкие особенности этих сортов обуславливают их устойчивость или чувствительность? Чтобы ответить на него, следует выявить маркеры устойчивости. Ими могут быть ферменты, связанные с устойчивостью, и цитогенетические особенности (частоты нарушений и митотический индекс). Если бы удалось обнаружить характерные особенности сортов на этих уровнях организации, можно было бы утверждать, что полиморфизм их устойчивости по морфологическим признакам не случаен, а связан с особенностями организации изученных растений на молекулярном и клеточном уровнях. В результате этих исследований появлялась возможность выделить биохимические маркеры, указывающие на высокую вероятность развития устойчивых или чувствительных реакций ячменя (и, возможно, других сельскохозяйственных растений) на действие техногенных стрессоров, в частности ТМ.
Вначале мы выделили белковые экстракты из зародышей семян контрастных по устойчивости сортов. Экстракты разделяли с помощью вертикального электрофореза в полиакриламидном геле с последующим специфическим окрашиванием на ферменты — супероксиддисмутазу (SOD), глутаматдегидрогеназу (GDH) и пероксидазу (PER). Здесь стоит оговориться, что причины формирования полиморфизма могут крыться и в других областях, однако, судя по литературным данным, анализ этих ферментов весьма перспективен. Метод электрофореза основан на разделении крупных заряженных молекул высокомолекулярных соединений в пористом геле под действием электрического поля. Выбор указанных ферментов объясняется их связью с устойчивостью к неблагоприятным факторам среды. SOD — антиокислительный фермент, встречающийся у всех живых организмов и ответственный за детоксикацию супероксидрадикала. PER — тоже антиокислительный фермент, но разрушающий перекись водорода. GDH — фермент азотистого обмена, связанный с устойчивостью к стрессам [24]. В результате выдвинутая гипотеза подтвердилась: были обнаружены два аллеля SOD и один PER, с большей вероятностью наблюдаемые у чувствительных сортов, и один аллель GDH, характерный для устойчивых сортов. Их и можно рассматривать как маркеры чувствительности и устойчивости соответственно.
Далее мы перешли к цитогенетическому анализу. Подсчитывали частоты цитогенетических нарушений, которые затем сравнивали для групп контрастных сортов. В этом и вышеописанном экспериментах обработка опытных образцов солями свинца не производилась. Такое решение было принято, чтобы установить, являются ли различия между контрастными сортами следствием внутренних (генетических) особенностей растений или они проявляются лишь под действием стрессора. Результатом этого блока исследований стало выявление маркеров, связанных с развитием устойчивых или чувствительных реакций растений на действие свинца — изоэнзимных и цитогенетических. Под первыми следует понимать специфические аллельные варианты, с большей вероятностью встречающиеся у той или иной группы сортов, а во втором — цитогенетические аномалии, чаще встречающиеся у представителей одной из групп, и разницу в других показателях, связанных с особенностями пролиферации. Эксперимент дал хорошие результаты: было обнаружено, что спонтанные частоты цитогенетических аномалий (многополюсные митозы, слипания хромосом, мосты) у чувствительных сортов достоверно выше, чем у устойчивых. То есть чувствительные сорта более склонны к мутациям.
Сделанные ранее выводы и полученные результаты вполне справедливы: действительно, существуют различия между сортами ячменя в плане их устойчивости к тяжелым металлам. Имеются сорта, которые достаточно успешно приспосабливаются к наличию в среде поллютанта и могут нормально завершать свой жизненный цикл, давая урожай. С другой стороны, есть и такие сорта, которые очень чувствительны к действию загрязнителя и фактически уже не могут существовать в таких условиях. В лучшем случае эти растения еще способны поддерживать свою жизнеспособность, но все физиологически значимые показатели у них будут сильно снижены. В худшем случае такие растения обречены на гибель. Поэтому в условиях реального сельскохозяйственного производства выращивание чувствительных сортов на загрязненных территориях будет убыточным. Хотя и не верно было бы совсем отказываться от них в реальной практике, потому что, как указывалось ранее, устойчивость может достигаться ухудшением иных жизненных показателей [9].
Мы показали, что обнаруженные различия вовсе не случайны и не зависят от внешних факторов. Устойчивость или чувствительность к воздействию тяжелых металлов определяется генетическими особенностями организма, которые можно считать сортовым признаком. Существуют определенные изоформы ферментов, более эффективные в плане защиты от внешних негативных агентов, позволяющие растениям успешнее противостоять стрессам. Те сорта, которые ими обладают, в среднем лучше приспосабливаются к неблагоприятным условиям. Более того, мы обнаружили и другие тонкие особенности контрастных сортов, определяющие развитие у них устойчивых или чувствительных реакций на действие свинца. Выяснилось, что для чувствительных сортов характерен более высокий выход цитогенетических нарушений по сравнению с тем, что наблюдается у устойчивых сортов. Это говорит о меньшей надежности хромосомного аппарата чувствительных сортов. Вероятно, в дальнейшем имеет смысл определить гены, контролирующие синтез этих изоформ, и использовать их для создания новых сортов ячменя. Это особенно актуально потому, что современные культурные растения существенно потеряли в генетическом разнообразии вследствие селекции на хозяйственно ценные признаки [25], и это негативно сказывается на их устойчивости. Для сельского хозяйства полезно подобрать оптимальные районированные сорта, пригодные для выращивания при актуальных уровнях техногенного загрязнения.
Можно отметить несколько вероятных направлений дальнейшего развития нашей работы. Первое — это оценка действия на уже использованные сорта ячменя других распространенных тяжелых металлов, например, кадмия. Будут ли воспроизводиться полученные результаты? Другое направление — поиск молекулярных маркеров устойчивости/чувствительности к действию ТМ у других культур. Характерны ли обнаруженные особенности только для ячменя? Помимо того, известны устойчивые сорта, которые способны успешно развиваться на обогащенной свинцом среде. Вызывает интерес вопрос о том, какое количество этого ТМ может попасть в товарную часть растения — зерно? Если окажется, что оно накапливает свинец в опасных для здоровья человека количествах, то это ограничит применение результатов.
Устойчивость к болезням растений — Plant disease resistance

На этой диаграмме показан процесс от прикрепления грибов или бактерий к растительной клетке до определенного типа реакции. PTI означает иммунитет, запускаемый образцом, а ETI — иммунитет, запускаемый эффектором .
Устойчивость растений к болезням защищает растения от патогенов двумя способами: с помощью заранее сформированных структур и химических веществ, а также с помощью реакций иммунной системы, вызванных инфекциями. По сравнению с восприимчивым растением устойчивость к болезням — это уменьшение роста патогенов на растении или внутри него (и, следовательно, уменьшение заболеваемости), в то время как термин « толерантность к болезням» описывает растения, которые проявляют небольшое повреждение болезнями, несмотря на значительные уровни патогенов. Исход болезни определяется трехсторонним взаимодействием патогена, растения и условий окружающей среды (взаимодействие, известное как треугольник болезни ).
Соединения, активирующие защиту, могут перемещаться от клетки к клетке и систематически через сосудистую систему растения. Однако у растений нет циркулирующих иммунных клеток, поэтому большинство типов клеток демонстрируют широкий набор противомикробных защитных механизмов . Хотя очевидные качественные различия в устойчивости к болезням могут наблюдаться при сравнении нескольких образцов (что позволяет классифицировать их как «устойчивые» или «чувствительные» после заражения одним и тем же штаммом патогена при аналогичных уровнях посевного материала в аналогичных средах), градация количественных различий в устойчивости к болезням чаще наблюдается между штаммами или генотипами растений . Растения постоянно сопротивляются одним патогенам, но уступают другим; устойчивость обычно специфична для определенных видов патогенов или штаммов патогенов.
СОДЕРЖАНИЕ
- 4.3.1 Убиквитин и передача сигналов E3
- 6.1 Устойчивость к ГМ или трансгенным инженерным болезням
Устойчивость растений к болезням имеет решающее значение для надежного производства продуктов питания и обеспечивает значительное сокращение сельскохозяйственного использования земли, воды, топлива и других ресурсов. Растения как в естественных, так и в культурных популяциях обладают естественной устойчивостью к болезням, но это не всегда защищает их.
Фитофтороз Великий голод в Ирландии 1840-х годов был вызван оомицетом Phytophthora infestans . Первый в мире сорт бананов массового выращивания Gros Michel был утерян в 1920-х годах из-за панамской болезни, вызванной грибком Fusarium oxysporum . Современные эпидемии стеблевой ржавчины пшеницы, листовой ржавчины и желтой полосатой ржавчины, распространяющиеся из Восточной Африки на Индийский субконтинент, вызываются ржавчинными грибами Puccinia graminis и P. striiformis . Другие эпидемии включают Честнат трущобы , а также рецидивирующие тяжелые заболевания растений, такие как Райс взрыва , соевой нематодой , Citrus язву .
Патогены растений могут быстро распространяться на большие расстояния под воздействием воды, ветра, насекомых и людей. Подсчитано, что в крупных регионах и для многих видов сельскохозяйственных культур болезни обычно снижают урожайность растений на 10% каждый год в более развитых странах или сельскохозяйственных системах, но потери урожая от болезней часто превышают 20% в менее развитых странах.
Однако борьба с болезнями является достаточно успешной для большинства сельскохозяйственных культур. Борьба с болезнями достигается за счет использования растений, которые были выведены с целью обеспечения хорошей устойчивости ко многим заболеваниям, и подходов к выращиванию растений, таких как севооборот, семена, свободные от патогенов, подходящая дата посадки и густота растений, контроль влажности поля и использование пестицидов .
Общие механизмы устойчивости к болезням
Готовые конструкции и компаунды

- Кутикула растения / поверхность
- Стенки растительных клеток
- Противомикробныехимические вещества (например: полифенолы, сесквитерпеновые лактоны, сапонины )
- Детоксифицирующие ферменты, расщепляющие токсины патогенного происхождения
- Рецепторы, которые воспринимают присутствие патогена и активируют индуцируемую защиту растений
Индуцируемая постинфекционная защита растений
- Армирование клеточной стенки ( целлюлоза, лигнин, суберин, каллоза, белки клеточной стенки)
- Противомикробные химические вещества, включая активные формы кислорода, такие как перекись водорода или пероксинитрит, или более сложные фитоалексины, такие как генистеин или камалексин.
- Антимикробные белки, такие как дефенсины, тионины или PR-1
- Антимикробные ферменты, такие как хитиназы, бета- глюканазы или пероксидазы
- Гиперчувствительный ответ — быстрый ответ смерти клетки-хозяина, связанный с индукцией защиты.
Иммунная система
Иммунная система растений несет два взаимосвязанных уровня рецепторов, один из которых чаще всего воспринимает молекулы вне клетки, а другой — молекулы внутри клетки. Обе системы обнаруживают злоумышленника и реагируют, активируя противомикробную защиту в инфицированной клетке и соседних клетках. В некоторых случаях сигналы активации защиты распространяются на остальную часть растения или даже на соседние растения. Обе системы обнаруживают различные типы молекул патогенов и классы белков рецепторов растений.
Первый уровень в первую очередь регулируется рецепторами распознавания образов, которые активируются распознаванием эволюционно консервативных патогенов или молекулярных паттернов, ассоциированных с микробами (PAMPs или MAMPs). Активация PRR приводит к внутриклеточной передаче сигналов, транскрипционному репрограммированию и биосинтезу сложной выходной реакции, которая ограничивает колонизацию. Система известна как иммунитет, запускаемый PAMP, или иммунитет, запускаемый образцом (PTI).
Второй уровень, в первую очередь регулируемый продуктами гена R, часто называют иммунитетом, запускаемым эффектором (ETI). ETI обычно активируется присутствием специфических «эффекторов» патогена, а затем запускает сильные противомикробные реакции (см. Раздел гена R ниже).
В дополнение к PTI и ETI, защита растений может быть активирована путем восприятия соединений, связанных с повреждением (DAMP), таких как участки стенки растительной клетки, высвобождаемые во время патогенной инфекции.
Ответы, активируемые рецепторами PTI и ETI, включают стробирование ионных каналов, окислительный взрыв, клеточные окислительно-восстановительные изменения или каскады протеинкиназ, которые непосредственно активируют клеточные изменения (например, усиление клеточной стенки или производство антимикробных препаратов) или активируют изменения в экспрессии генов, которые затем усиливают другие защитные механизмы. ответы
Иммунная система растений демонстрирует некоторые механистические сходства с иммунными системами насекомых и млекопитающих, но также демонстрирует многие специфические для растений характеристики. Два вышеописанных уровня являются центральными для иммунитета растений, но не полностью описывают иммунную систему растений. Кроме того, многие конкретные примеры очевидных PTI или ETI нарушают общие определения PTI / ETI, что указывает на необходимость расширенных определений и / или парадигм.
Иммунитет, вызываемый паттерном
PAMP, консервативные молекулы, которые населяют несколько родов патогенов, многие исследователи называют MAMP. Защиты, вызванные восприятием MAMP, достаточны для отражения большинства патогенов. Однако эффекторные белки патогенов (см. Ниже) адаптированы для подавления базальной защиты, такой как PTI. Обнаружено множество рецепторов MAMP (и DAMP). MAMP и DAMP часто обнаруживаются трансмембранными рецепторными киназами, которые несут внеклеточные домены LRR или LysM .
Эффекторный иммунитет
Иммунитет, запускаемый эффектором (ETI), активируется присутствием эффекторов патогена. Ответ ETI зависит от генов R и активируется конкретными штаммами патогенов. ETI растений часто вызывает гиперчувствительный апоптотический ответ .
Гены R и белки R
В растениях развились гены R ( гены устойчивости), продукты которых обеспечивают устойчивость к конкретным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты гена R представляют собой белки, которые позволяют распознавать эффекторы конкретных патогенов либо путем прямого связывания, либо путем распознавания изменения эффектора белка-хозяина. Многие гены R кодируют белки NB-LRR (белки с нуклеотид-связывающими и богатыми лейцином повторяющимися доменами, также известные как белки NLR или белки STAND, среди других названий). Большинство иммунных систем растений несут в себе репертуар из 100-600 различных гомологов R-гена. Было продемонстрировано, что отдельные гены R опосредуют устойчивость к конкретным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты гена R контролируют широкий набор реакций устойчивости к болезням, индукции которых часто бывает достаточно, чтобы остановить дальнейший рост / распространение патогенов.
Изученные гены R обычно придают специфичность конкретным штаммам видов патогенов (те, которые экспрессируют признанный эффектор). Как впервые отметил Гарольд Флор в его формулировке отношения ген-ген в середине 20-го века, ген R растения имеет специфичность в отношении гена авирулентности патогена (гена Avr). В настоящее время известно, что гены авирулентности кодируют эффекторы. Ген патогена Avr должен обладать такой же специфичностью, что и ген R, чтобы этот ген R придавал устойчивость, что предполагает взаимодействие рецептор / лиганд для генов Avr и R. В качестве альтернативы эффектор может модифицировать свою клеточную мишень-хозяин (или молекулярную ловушку этой мишени), а продукт гена R (белок NLR) активирует защиту, когда он обнаруживает модифицированную форму мишени-хозяина или приманки.
Эффекторная биология
Эффекторы играют центральную роль в патогенном или симбиотическом потенциале микробов и микроскопических животных, колонизирующих растения, таких как нематоды. Эффекторы обычно представляют собой белки, которые доставляются за пределы микроба в клетку-хозяина. Эти производные от колонистов эффекторы влияют на физиологию и развитие клетки-хозяина. Таким образом, эффекторы представляют собой примеры совместной эволюции (пример: грибковый белок, который функционирует вне гриба, но внутри растительных клеток, эволюционировал, чтобы взять на себя специфические для растения функции). Диапазон хозяев патогена определяется, среди прочего, наличием соответствующих эффекторов, которые делают возможной колонизацию конкретного хозяина. Эффекторы, происходящие от патогенов, являются мощным инструментом для определения функций растений, которые играют ключевую роль в развитии болезней и устойчивости к ним. По-видимому, большинство эффекторов функционируют, чтобы манипулировать физиологией хозяина, чтобы вызвать заболевание. Хорошо изученные бактериальные патогены растений обычно экспрессируют несколько десятков эффекторов, которые часто доставляются хозяину с помощью аппарата секреции типа III . Патогены грибов, оомицетов и нематод, по-видимому, экспрессируют несколько сотен эффекторов.
Так называемые «основные» эффекторы определяются оперативно по их широкому распространению в популяции конкретного патогена и их значительному вкладу в вирулентность патогена. Геномика может быть использована для идентификации основных эффекторов, которые затем могут быть использованы для обнаружения новых аллелей гена R, которые могут быть использованы в селекции растений на устойчивость к болезням.
Малые РНК и РНК-интерференция
Подразумевается, что пути растительных мРНК являются важными компонентами иммунитета, запускаемого патоген-ассоциированным молекулярным паттерном (PAMP), и иммунитета, запускаемого эффектором (ETI). Было показано, что индуцированные бактериями миРНК у Arabidopsis влияют на гормональную передачу сигналов, включая ауксин, абсцизовую кислоту (ABA), жасмоновую кислоту (JA) и салициловую кислоту (SA). Успехи полногеномных исследований выявили массовую адаптацию паттернов экспрессии миРНК хозяина после заражения грибковыми патогенами Fusarium virguliforme, Erysiphe graminis, Verticillium dahliae и Cronartium quercuum, а также оомицетом Phytophthora sojae . Изменения экспрессии мРНК в ответ на грибковые патогены указывают на то, что в этом пути защиты может быть задействовано молчание генов. Однако есть свидетельства того, что противогрибковая защитная реакция на Colletotrichum spp. Инфекция кукурузы не полностью регулируется специфической индукцией miRNA, но вместо этого может действовать для точной настройки баланса между генетическими и метаболическими компонентами при заражении.
Транспорту мРНК во время инфекции, вероятно, способствуют внеклеточные везикулы (EV) и мультивезикулярные тельца (MVB). Состав РНК в растительных EV полностью не изучен, но вполне вероятно, что они частично ответственны за транспортировку РНК. Растения могут транспортировать вирусные РНК, мРНК, микроРНК (миРНК) и малые интерферирующие РНК (миРНК) системно через флоэму. Считается, что этот процесс происходит через плазмодесмы и включает РНК-связывающие белки, которые способствуют локализации РНК в клетках мезофилла. Хотя они были идентифицированы во флоэме с помощью мРНК, нет убедительных доказательств того, что они опосредуют транспорт РНК на большие расстояния. Следовательно, EV могут вносить вклад в альтернативный путь загрузки РНК во флоэму или, возможно, могут транспортировать РНК через апопласт. Есть также свидетельства того, что растительные EV могут обеспечивать межвидовой перенос мРНК посредством РНК-интерференции, такой как генное молчание, индуцированное хозяином (HIGS). Транспорт РНК между растениями и грибами, по-видимому, является двунаправленным, поскольку было показано, что мРНК от грибкового патогена Botrytis cinerea нацелены на гены защиты хозяина у арабидопсиса и томатов.
Сопротивление на уровне видов
В небольшом количестве случаев гены растений эффективны против целого вида патогена, даже если этот вид является патогенным для других генотипов этого вида хозяина. Примеры включают MLO ячменя против мучнистой росы, пшеницу Lr34 против листовой ржавчины и пшеницу Yr36 против полосовой ржавчины пшеницы . Ряд механизмов этого типа устойчивости может существовать в зависимости от конкретного гена и комбинации растение-патоген. Другие причины эффективного иммунитета растений могут включать в себя отсутствие коадаптации (патоген и / или растение лишены множества механизмов, необходимых для колонизации и роста в пределах этого вида-хозяина) или особенно эффективный набор заранее сформированных защитных механизмов.
Механизмы сигнализации
Восприятие присутствия патогена
Передача сигналов защиты растений активируется рецепторами, обнаруживающими патоген, которые описаны в предыдущем разделе. Активированные рецепторы часто вызывают образование реактивного кислорода и оксида азота, потоки ионов кальция, калия и протонов , измененные уровни салициловой кислоты и других гормонов и активацию киназ MAP и других специфических протеинкиназ . Эти события, в свою очередь, обычно приводят к модификации белков, контролирующих транскрипцию генов , и активации экспрессии генов, связанных с защитой .
Факторы транскрипции и гормональный ответ
Были идентифицированы многочисленные гены и / или белки, а также другие молекулы, которые опосредуют передачу защитного сигнала растений. Динамика движения цитоскелета и пузырьков помогает ориентировать защитные реакции растений на точку атаки патогена.
Механизмы факторов транскрипции и гормонов
Активность иммунной системы растений частично регулируется сигнальными гормонами, такими как:
Между этими путями может существовать существенная перекрестная связь.
Регулирование деградацией
Как и во многих путях передачи сигналов, экспрессия генов растений во время иммунных ответов может регулироваться деградацией. Это часто происходит, когда связывание гормона с рецепторами гормонов стимулирует ассоциированную с убиквитином деградацию репрессорных белков, которые блокируют экспрессию определенных генов. Конечный результат — экспрессия генов, активируемых гормонами. Примеры:
- Ауксин : связывается с рецепторами, которые затем рекрутируют и разрушают репрессоры активаторов транскрипции, которые стимулируют экспрессию ауксин-специфических генов.
- Жасмоновая кислота: аналогична ауксину, за исключением рецепторов жасмоната, влияющих на сигнальные медиаторы ответа жасмоната, такие как белки JAZ.
- Гибберелловая кислота : Гиббереллин вызывает конформационные изменения рецептора, а также связывание и деградацию белков Della .
- Этилен: ингибирующее фосфорилирование активатора реакции этилена EIN2 блокируется связыванием этилена. Когда это фосфорилирование снижается, белок EIN2 расщепляется, и часть белка перемещается в ядро, чтобы активировать экспрессию гена ответа на этилен.
Убиквитин и передача сигналов E3
Убиквитинирование играет центральную роль в передаче клеточных сигналов, которая регулирует процессы, включая деградацию белка и иммунологический ответ. Хотя одной из основных функций убиквитина является нацеливание на белки для разрушения, он также полезен в сигнальных путях, высвобождении гормонов, апоптозе и перемещении материалов по клетке. Убиквитинирование — это компонент нескольких иммунных ответов. Без правильного функционирования убиквитина вторжение патогенов и других вредных молекул резко увеличится из-за ослабленной иммунной защиты.
E3 сигнализация
Фермент убиквитинлигаза E3 является основным компонентом, который обеспечивает специфичность путей деградации белка, включая пути передачи сигналов иммунной системы. Компоненты фермента E3 могут быть сгруппированы по доменам, которые они содержат, и включают несколько типов. К ним относятся одиночный подблок Ring и U-box, HECT и CRL . Сигнальные пути растений, включая иммунные ответы, контролируются несколькими путями обратной связи, которые часто включают отрицательную обратную связь; и они могут регулироваться ферментами деубиквитинирования, деградацией факторов транскрипции и деградацией негативных регуляторов транскрипции.

На этом изображении показаны пути, проистекающие из реакции иммунитета растений. Это подчеркивает роль и эффект убиквитина в регулировании этого пути.
Сроки иммуногенеза
Устойчивость рассады
Устойчивость проростков — это сопротивление, которое начинается на стадии развития растения и сохраняется на протяжении всей его жизни. Он не предназначен для понимания сопротивления, которое проявляется только на стадии прорастания. Синоним основного гена устойчивости .
Устойчивость взрослых растений
Устойчивость взрослых растений не проявляется на стадии проростков, но существует на протяжении остальной части жизни растения. Разница между устойчивостью проростков и устойчивостью взрослых особей особенно важна для однолетних культур . APR и количественное сопротивление часто идут вместе, хотя они не являются синонимами. Синоним минорного гена устойчивости .
Селекция растений на устойчивость к болезням
Селекционеры уделяют особое внимание отбору и развитию устойчивых к болезням линий растений. Болезни растений также можно частично контролировать с помощью пестицидов и таких методов выращивания, как севооборот, обработка почвы, плотность посадки, семена без болезней и очистка оборудования, но обычно предпочтительны сорта растений с присущей (генетически детерминированной) устойчивостью к болезням. Селекция на устойчивость к болезням началась, когда растения были впервые одомашнены. Усилия по селекции продолжаются, потому что популяции патогенов находятся под давлением отбора из- за повышенной вирулентности, появляются новые патогены, развивающиеся методы выращивания и изменение климата могут снизить устойчивость и / или усилить патогены, а селекция растений по другим признакам может нарушить предшествующую устойчивость. Линия растений с приемлемой устойчивостью к одному патогену может не иметь устойчивости к другим.
Селекция на устойчивость обычно включает:
- Идентификация растений, которые могут быть менее желательными в других отношениях, но которые несут полезный признак устойчивости к болезням, включая линии диких растений, которые часто проявляют повышенную устойчивость.
- Скрещивание желаемого, но восприимчивого к болезням сорта с растением, которое является источником устойчивости.
- Рост селекционных кандидатов в благоприятных для болезни условиях, возможно, включая прививку патогенов. Необходимо уделять внимание изолятам конкретных патогенов, чтобы решить проблему изменчивости в пределах одного вида патогенов.
- Отбор устойчивых к болезням особей, которые сохраняют другие желательные характеристики, такие как урожайность, качество, а также другие характеристики устойчивости к болезням.
Устойчивость называется устойчивой, если она продолжает оставаться эффективной в течение многих лет широкого использования по мере развития популяций патогенов. « Вертикальная устойчивость » специфична для определенных рас или штаммов видов патогенов, часто контролируется отдельными генами R и может быть менее устойчивой. Горизонтальная устойчивость или устойчивость широкого спектра против целого вида патогенов часто бывает не полностью эффективной, но более устойчивой и часто контролируется многими генами, которые сегрегируют в племенных популяциях. Устойчивость устойчивости важна даже тогда, когда ожидается, что в будущем появятся улучшенные сорта: среднее время от распознавания человеком новой угрозы грибкового заболевания до выпуска устойчивой культуры для этого патогена составляет не менее двенадцати лет.
Такие культуры, как картофель, яблоки, бананы и сахарный тростник, часто размножаются путем вегетативного размножения для сохранения очень желательных сортов растений, потому что для этих видов ауткроссинг серьезно нарушает предпочтительные признаки. См. Также бесполое размножение . Вегетативно размножаемые культуры могут быть одной из лучших целей для повышения устойчивости с помощью биотехнологического метода трансформации растений для управления генами, влияющими на устойчивость к болезням.
Научная селекция устойчивости к болезням началась с сэра Роуленда Биффена , который идентифицировал единственный рецессивный ген устойчивости к желтой ржавчине пшеницы. Затем почти все культуры были выведены с включением генов устойчивости к болезням (R), многие из которых были интрогрессированы от совместимых диких родственников.
Устойчивость к ГМ или трансгенным инженерным болезням
Термин GM ( «генетически модифицированный» ) часто используется как синоним трансгенного для обозначения растений, модифицированных с использованием технологий рекомбинантной ДНК. Растения с устойчивостью к трансгенным / ГМ болезням против насекомых-вредителей оказались чрезвычайно успешными в качестве коммерческих продуктов, особенно кукурузы и хлопка, и ежегодно их высаживают на площади более 20 миллионов гектаров в более чем 20 странах мира (см. Также генетически модифицированные культуры ). Устойчивость трансгенных растений к болезням против микробных патогенов была впервые продемонстрирована в 1986 году. Экспрессия последовательностей генов белков оболочки вируса придает устойчивость к вирусу посредством малых РНК . Это оказалось широко применимым механизмом для подавления репликации вирусов. Объединив гены белков оболочки от трех разных вирусов, ученые разработали гибриды кабачков с подтвержденной полевирусной устойчивостью. Подобные уровни устойчивости к этому разнообразию вирусов не были достигнуты с помощью обычного разведения.
Аналогичная стратегия была развернута в борьбу с папайей вируса кольцевой пятнистости, который к 1994 году под угрозу уничтожить Гавайи «s папайи промышленность. Полевые испытания показали отличную эффективность и высокое качество плодов. К 1998 году первая трансгенная папайя, устойчивая к вирусам, была одобрена для продажи. Устойчивость к болезням сохраняется более 15 лет. Трансгенная папайя составляет ~ 85% производства на Гавайях. Фрукт разрешен к продаже в США, Канаде и Японии.
Линии картофеля, экспрессирующие последовательности вирусных репликаз, которые придают устойчивость к вирусу скручивания листьев картофеля, продавались под торговыми наименованиями NewLeaf Y и NewLeaf Plus и широко применялись в коммерческом производстве в 1999-2001 годах, пока McDonald’s Corp. не решила не покупать ГМ-картофель, и компания Monsanto решила закрыть свой картофельный бизнес в NatureMark. Картофель NewLeaf Y и NewLeaf Plus несут два ГМ-признака, поскольку они также выражают Bt-опосредованную устойчивость к колорадскому жуку.
К 2013 году на рынке не было ни одной другой культуры с искусственно созданной устойчивостью к болезням против патогенных микроорганизмов, хотя более десятка культур находились в стадии разработки и тестирования.
| Год публикации | Обрезать | Устойчивость к болезням | Механизм | Статус разработки |
|---|---|---|---|---|
| 2012 г. | Помидор | Бактериальное пятно | Ген R из перца | 8 лет полевых испытаний |
| 2012 г. | Рис | Бактериальный ожог и бактериальная полоса | Разработанный ген E | Лаборатория |
| 2012 г. | Пшеница | Мучнистая роса | Сверхэкспрессия гена R из пшеницы | 2 года полевых испытаний на момент публикации |
| 2011 г. | яблоко | Грибок парши яблони | Ген тионина из ячменя | 4 года полевых испытаний на момент публикации |
| 2011 г. | Картошка | Картофельный вирус Y | Резистентность, вызванная патогенами | 1 год полевых испытаний на момент публикации |
| 2010 г. | яблоко | Бактериальный ожог | Антибактериальный протеин от моли | 12 лет полевых испытаний на момент публикации |
| 2010 г. | Помидор | Мультибактериальная устойчивость | PRR от Arabidopsis | Лабораторные весы |
| 2010 г. | Банан | Xanthomonas увядание | Новый ген из перца | Сейчас в полевых испытаниях |
| 2009 г. | Картошка | Фитофтороз | Гены R от диких родственников | 3 года полевых испытаний |
| 2009 г. | Картошка | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 г. | Картошка | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 г. | слива | Вирус оспы сливы | Резистентность, вызванная патогенами | Нормативные разрешения, коммерческие продажи отсутствуют |
| 2005 г. | Рис | Бактериальная полоса | Ген R из кукурузы | Лаборатория |
| 2002 г. | Ячмень | Стеблевая ржавчина | Ген покоящейся лимфоцитарной киназы (RLK) из устойчивого сорта ячменя | Лаборатория |
| 1997 г. | Папайя | Вирус кольцевой пятнистости | Резистентность, вызванная патогенами | Одобрено и продается на коммерческой основе с 1998 года, продается в Японии с 2012 года. |
| 1995 г. | Давить | Три вируса мозаики | Резистентность, вызванная патогенами | Одобрено и продается с 1994 года. |
| 1993 г. | Картошка | Картофельный вирус X | Фермент, индуцированный интерфероном млекопитающих | 3 года полевых испытаний на момент публикации |
Перенос PRR
Исследования, направленные на искусственное сопротивление, основываются на нескольких стратегиях. Один из них — передать полезные PRR видам, у которых они отсутствуют. Идентификация функциональных PRR и их передача реципиентному виду, у которого отсутствует ортологичный рецептор, может обеспечить общий путь к дополнительным расширенным репертуарам PRR. Например, рецептор EF-Tu (EFR) PRR арабидопсиса распознает бактериальный фактор элонгации трансляции EF-Tu . Исследования, проведенные в лаборатории Сейнсбери, показали, что использование EFR в Nicotiana benthamiana или Solanum lycopersicum (томат), которые не могут распознавать EF-Tu , придает устойчивость к широкому спектру бактериальных патогенов. Экспрессия EFR в томатах была особенно эффективной против широко распространенной и разрушительной почвенной бактерии Ralstonia solanacearum . И наоборот, ген PRR Verticillium 1 ( Ve1 ) томата может быть перенесен от томата к Arabidopsis , где он придает устойчивость изолятам Verticillium расы 1 .
Штабелирование
Вторая стратегия пытается задействовать несколько генов NLR одновременно, стратегия разведения известна как стекинг. Сорта, полученные путем молекулярной селекции с помощью ДНК или переноса генов, вероятно, будут проявлять более устойчивую устойчивость, потому что патогены должны будут мутировать несколько эффекторных генов. Секвенирование ДНК позволяет исследователям функционально «добывать» гены NLR из нескольких видов / штаммов.
AvrBs2 эффекторный ген из Xanthomona огапза является возбудителем бактериальной пятнистости перца и томатов. Первый «эффекторно-рациональный» поиск потенциально долговечного гена R последовал за открытием, что avrBs2 обнаруживается у большинства видов Xanthomonas, вызывающих болезни, и необходим для приспособленности к патогенам. Ген Bs2 NLR из дикого перца, Capsicum chacoense, был перенесен в томат, где он ингибировал рост патогена. Полевые испытания продемонстрировали высокую устойчивость без бактерицидных химикатов. Однако редкие штаммы Xanthomonas преодолели Bs2-опосредованную устойчивость перца путем приобретения мутаций avrBs2, которые избегают распознавания, но сохраняют вирулентность. Сложение генов R, каждый из которых распознает разные основные эффекторы, может задерживать или предотвращать адаптацию.
Более 50 локусов в штаммах пшеницы придают устойчивость к болезням против возбудителей стеблевой, листовой и желтой полосатой ржавчины пшеницы. Ген NLR Stem rust 35 ( Sr35 ), клонированный от диплоидного родственника культурной пшеницы, Triticum monococcum, обеспечивает устойчивость к изоляту пшеничной ржавчины Ug99 . Сходным образом, Sr33, происходящий от пшеничного родственника Aegilops tauschii, кодирует пшеничный ортолог генов устойчивости к мучнистой росе Mla ячменя . Оба гена необычны для пшеницы и ее родственников. В сочетании с геном Sr2, который действует аддитивно, по крайней мере, с Sr33, они могут обеспечить прочную устойчивость к болезням к Ug99 и его производным.
Гены-исполнители
Другой класс генов устойчивости растений к болезням открывает «лазейку», которая быстро убивает пораженные клетки, останавливая размножение патогенов. Эффекторы, подобные активаторам транскрипции (TAL) Xanthomonas и Ralstonia, представляют собой ДНК-связывающие белки, которые активируют экспрессию генов хозяина для повышения вирулентности патогенов. И линии риса, и перца независимо развили сайты связывания TAL-эффекторов, которые вместо этого действуют как «палач», вызывающий гибель гиперчувствительных клеток-хозяев при активации. Xa27 из риса и Bs3 и Bs4c из перца являются такими генами «исполнителей» (или «исполнителей»), которые кодируют негомологичные растительные белки с неизвестной функцией. Гены-исполнители экспрессируются только в присутствии определенного эффектора TAL.
Сконструированные гены-исполнители были продемонстрированы путем успешного изменения конструкции промотора Bs3 перца, чтобы он содержал два дополнительных сайта связывания для эффекторов TAL из разнородных штаммов патогенов. Впоследствии сконструированный ген-исполнитель был внедрен в рис путем добавления пяти эффекторных сайтов связывания TAL к промотору Xa27 . Синтетическая конструкция Xa27 придает устойчивость к бактериальному ожогу Xanthomonas и видам бактериальных полос на листьях.
Аллели восприимчивости хозяина
Большинство патогенов растений перепрограммируют паттерны экспрессии генов-хозяев, чтобы непосредственно принести пользу патогену. Перепрограммированные гены, необходимые для выживания и размножения патогенов, можно рассматривать как «гены восприимчивости к болезням». Рецессивные гены устойчивости являются кандидатами на предрасположенность к болезням. Например, мутация отключила ген Arabidopsis, кодирующий пектатлиазу (участвующую в деградации клеточной стенки), придавая устойчивость возбудителю мучнистой росы Golovinomyces cichoracearum . Точно так же ген MLO ячменя и спонтанно мутировавшие ортологи MLO гороха и томата также придают устойчивость к мучнистой росе.
Lr34 — это ген, обеспечивающий частичную устойчивость к листовой и желтой ржавчине и мучнистой росе пшеницы. Lr34 кодирует переносчик аденозинтрифосфата (АТФ) -связывающей кассеты (ABC). Доминантный аллель, обеспечивающий устойчивость к болезням, недавно был обнаружен у культурной пшеницы (не у диких штаммов) и, как и MLO, обеспечивает устойчивость широкого спектра у ячменя.
Природные аллели факторов инициации элонгации трансляции хозяина eif4e и eif4g также являются рецессивными генами вирусной устойчивости. Некоторые из них были применены для борьбы с потивирусами в ячмене, рисе, помидорах, перце, горохе, салате и дыне. Это открытие привело к успешному скринингу мутантов на химически индуцированные аллели eif4e в томате.
Естественная вариация промотора может привести к развитию рецессивных аллелей устойчивости к болезням. Например, рецессивный ген устойчивости xa13 в рисе является аллелем Os-8N3 . Os-8N3 транскрипционно активируется Xanthomonas oryzae pv. oryzae, экспрессирующие эффекторный TAL PthXo1 . Ген xa13 имеет в своем промоторе мутантный эффектор-связывающий элемент, который устраняет связывание PthXo1 и делает эти линии устойчивыми к штаммам, которые зависят от PthXo1 . Это открытие также продемонстрировало, что Os-8N3 необходим для восприимчивости.
Xa13 / Os-8N3 необходим для развития пыльцы, показывая, что такие мутантные аллели могут быть проблематичными, если фенотип восприимчивости к заболеванию изменяет функцию в других процессах. Однако мутации в эффекторно-связывающем элементе Os11N3 (OsSWEET14) TAL были произведены путем слияния эффекторов TAL с нуклеазами ( TALENs ). Отредактированные геномом растения риса с измененными сайтами связывания Os11N3 оставались устойчивыми к Xanthomonas oryzae pv. oryzae, но все же обеспечивали нормальную функцию развития.
Подавление гена
Устойчивость, основанная на подавлении РНК, является мощным инструментом для создания устойчивых культур. Преимущество РНКи как новой генной терапии против грибковых, вирусных и бактериальных инфекций растений заключается в том, что она регулирует экспрессию генов посредством деградации информационной РНК, репрессии трансляции и ремоделирования хроматина с помощью небольших некодирующих РНК. Механически процессы сайленсинга управляются продуктами процессинга двухцепочечной РНК ( дцРНК ) триггера, которые известны как малые интерферирующие РНК и микроРНК .
Диапазон хостов
Среди тысяч видов патогенных микроорганизмов растений лишь незначительное меньшинство способно инфицировать широкий спектр видов растений. Вместо этого большинство патогенов проявляют высокую степень специфичности к хозяину. Часто говорят, что виды растений, не являющиеся хозяевами, проявляют устойчивость, не являющуюся хозяином . Термин « устойчивость к хозяину» используется, когда вид патогена может быть патогенным для вида-хозяина, но определенные штаммы этого вида растений сопротивляются определенным штаммам данного вида патогена. Причины резистентности хозяина и нехозяина могут совпадать. Диапазон хозяев патогена определяется, среди прочего, наличием соответствующих эффекторов, которые делают возможной колонизацию конкретного хозяина. Диапазон патогенов-хозяев может измениться довольно внезапно, если, например, способность патогена синтезировать специфический для хозяина токсин или эффектор достигается перетасовкой / мутацией генов или горизонтальным переносом генов .
Эпидемии и популяционная биология
Коренные популяции часто характеризуются значительным разнообразием генотипов и рассредоточенными популяциями (рост в смеси со многими другими видами растений). Они также претерпели совместную эволюцию патогенов с растениями . Следовательно, до тех пор, пока новые патогены не вводятся / не эволюционируют, такие популяции обычно демонстрируют лишь низкую частоту эпидемий тяжелых заболеваний .
Агротехнические системы с монокультурами обеспечивают идеальную среду для эволюции патогенов, поскольку они предлагают высокую плотность целевых образцов со схожими / идентичными генотипами. Рост мобильности, обусловленный современными транспортными системами, обеспечивает патогенам доступ к большему количеству потенциальных целей. Изменение климата может изменить жизнеспособный географический диапазон видов патогенных микроорганизмов и привести к тому, что некоторые болезни станут проблемой в районах, где ранее болезнь была менее значимой.
Эти факторы делают современное сельское хозяйство более подверженным эпидемиям болезней. Общие решения включают постоянную селекцию на устойчивость к болезням, использование пестицидов, использование пограничных инспекций и ограничений на импорт растений, поддержание значительного генетического разнообразия в генофонде сельскохозяйственных культур (см. Разнообразие культур ) и постоянный надзор для ускорения принятия соответствующих ответных мер. Некоторые виды патогенов обладают гораздо большей способностью преодолевать устойчивость растений к болезням, чем другие, часто из-за их способности быстро развиваться и широко распространяться.
Источник https://givoyles.ru/articles/uhod/ustoichivost-k-boleznyam/
Источник https://biomolecula.ru/articles/ustoichivost-rastenii-ot-molekul-i-genov-k-organizmu
Источник https://ru.abcdef.wiki/wiki/Plant_disease_resistance



